Биомеханика вдоха и выдоха физиология. Внешнее дыхание
3. Выдох. Биомеханизм выдоха. Процесс выдоха. Как происходит выдох?
4. Изменение объема легких во время вдоха и выдоха. Функция внутриплеврального давления. Плевральное пространство. Пневмоторакс.
5. Фазы дыхания. Объем легкого (легких). Частота дыхания. Глубина дыхания. Легочные объемы воздуха. Дыхательный объем. Резервный, остаточный объем. Емкость легких.
6. Факторы, влияющие на легочный объем в фазу вдоха. Растяжимость легких (легочной ткани). Гистерезис.
7. Альвеолы. Сурфактант. Поверхностное натяжение слоя жидкости в альвеолах. Закон Лапласа.
8. Сопротивление дыхательных путей. Сопротивление легких. Воздушный поток. Ламинарный поток. Турбулентный поток.
9. Зависимость «поток-объем» в легких. Давление в дыхательных путях при выдохе.
10. Работа дыхательных мышц в течение дыхательного цикла. Работа дыхательных мышц при глубоком дыхании.
Газообмен между атмосферным воздухом и альвеолярным пространством легких происходит в результате циклических изменений объема легких в течение фаз дыхательного цикла . В фазу вдоха объем легких увеличивается, воздух из внешней среды поступает в дыхательные пути и затем достигает альвеол. Напротив, в фазу выдоха происходит уменьшение объема легких и воздух из альвеол через дыхательные пути выходит во внешнюю среду. Увеличение и уменьшение объема легких обусловлены биомеханическими процессами изменения объема грудной полости при вдохе и выдохе.
Биомеханика дыхания. Биомеханика вдоха.
Рис. 10.1. Влияние сокращения диафрагмальной мышцы на объем грудной полости . Сокращение диафрагмальной мышцы при вдохе (пунктирная линия) вызывает опускание диафрагмы вниз, смещение органов брюшной полости вниз и вперед. В результате увеличивается объем грудной полости.Увеличение объема грудной полости при вдохе происходит в результате сокращения инспираторных мышц: диафрагмы и наружных межреберных. Основной дыхательной мышцей является диафрагма, которая находится в нижней трети грудной полости и разделяет грудную и брюшную полости. При сокращении диафрагмальной мышцы диафрагма движется вниз и смещает органы брюшной полости вниз и кпереди, увеличивая объем грудной полости преимущественно по вертикали (рис. 10.1).

Увеличению объема грудной полости при вдохе способствует сокращение наружных межреберных мышц, которые поднимают грудную клетку вверх, увеличивая объем грудной полости. Этот эффект сокращения наружных межреберных мышц обусловлен особенностями прикрепления мышечных волокон к ребрам - волокна идут сверху вниз и сзади кпереди (рис. 10.2). При подобном направлении мышечных волокон наружных межреберных мышц их сокращение поворачивает каждое ребро вокруг оси, проходящей через точки сочленения головки ребра с телом и поперечным отростком позвонка. В результате этого движения каждая нижележащая реберная дуга поднимается вверх больше, чем опускается вышерасположенная. Одновременное движение вверх всех реберных дуг приводит к тому, что грудина поднимается вверх и кпереди, а объем грудной клетки увеличивается в сагиттальной и фронтальной плоскостях. Сокращение наружных межреберных мышц не только увеличивает объем грудной полости, но и препятствует опусканию грудной клетки вниз. Например, у детей, имеющих неразвитые межреберные мышцы, грудная клетка уменьшается в размере во время сокращения диафрагмы (парадоксальное движение).
 Рис. 10.2. Направление волокон наружных межреберных мышц и увеличение объема грудной полости при вдохе
.
а - сокращение наружных межреберных мышц при вдохе поднимает нижнее ребро больше, чем опускает вниз верхнее. В результате реберные дуги поднимаются вверх и увеличивают (б) объем грудной полости в сагиттальной и фронтальной плоскости.
Рис. 10.2. Направление волокон наружных межреберных мышц и увеличение объема грудной полости при вдохе
.
а - сокращение наружных межреберных мышц при вдохе поднимает нижнее ребро больше, чем опускает вниз верхнее. В результате реберные дуги поднимаются вверх и увеличивают (б) объем грудной полости в сагиттальной и фронтальной плоскости.
При глубоком дыхании в биомеханизме вдоха , как правило, участвует вспомогательная дыхательная мускулатура - грудино-ключично-сосцевидные и передние лестничные мышцы, и их сокращение дополнительно увеличивает объем грудной клетки. В частности, лестничные мышцы поднимают верхние два ребра, а грудино-ключично-сосцевидные - поднимают грудину. Вдох является активным процессом и требует расхода энергии при сокращении инспираторных мышц, которая затрачивается на преодоление эластического сопротивления относительно ригидных тканей грудной клетки, эластического сопротивления легко растяжимой легочной ткани, аэродинамического сопротивления дыхательных путей потоку воздуха, а также на повышение внутриабдоминального давления и возникающего при этом смещения органов брюшной полости книзу.
Дыхание
Дыхание является одной из жизненно важных функций организма, направленной на поддержание оптимального уровня окислительно-восстановительных процессов в клетках. Дыхание - сложный биологический процесс, который обеспечивает доставку кислорода тканям, использование его клетками в процессе метаболизма и удаление образовавшегося углекислого газа.
Весь сложный процесс дыхания можно разделить на три основных этапа: внешнее дыхание, транспорт газов кровью и тканевое дыхание.
Внешнее дыхание - газообмен между организмом и окружающим его атмосферным воздухом. Внешнее дыхание в свою очередь можно разделить на два этапа:
Обмен газов между атмосферным и альвеолярным воздухом;
Газообмен между кровью легочных капилляров и альвеолярный воздухом (обмен газов в легких).
Транспорт газов кровью. Кислород и углекислый газ в свободном растворенном состоянии переносятся в незначительном количестве, основной объем этих газов транспортируется в связанном состоянии. Основным переносчиком кислорода является гемоглобин. С помощью гемоглобина транспортируется также до 20% углекислого газа (карбгемоглобин). Остальная часть углекислого газа переносится в виде бикарбонатов плазмы крови.
Внутреннее или тканевое дыхание. Этот этап дыхания можно разделить на два:
Обмен газов между кровью и тканями;
Потребление клетками кислорода и выделение углекислого газа.
Внешнее дыхание осуществляется циклически и состоит из фазы вдоха, выдоха и дыхательной паузы. У человека частота дыхательных движений в среднем равна 16-18 в одну минуту.
Биомеханика вдоха и выдоха
Вдох начинается с сокращения дыхательных (респираторных) мышц.
Мышцы, сокращение которых приводит к увеличению объема грудной полости, называются инспираторными (наружные межреберные, мышечные волокна диафрагмы, вспомогательные-грудинные, лестничные, грудинно-ключично-сосцевидные), а мышцы, сокращение которых приводит к уменьшению объема грудной полости, называются экспираторными (внутренние межреберные, вспомогательные – мышцы живота). Основной инспираторной мышцей является мышца диафрагмы. Сокращение мышцы диафрагмы приводит к тому, что купол ее уплощается, внутренние органы оттесняются вниз, что приводит к увеличению объема грудной полости в вертикальном направлении. Сокращение наружных межреберных и межхрящевых мышц приводит к увеличению объема грудной полости в сагитальном и фронтальном направлениях.
Легкие покрыты серозной оболочкой - плеврой, состоящей из висцерального и париетального листков. Париетальный листок соединен с грудной клеткой, а висцеральный - с тканью легких. При увеличении объема грудной клетки, в результате сокращения инспираторных мышц, париетальный листок последует за грудной клеткой. В результате появления адгезивных сил между листками плевры, висцеральный листок последует за париетальным, а вслед за ними и легкие. Это приводит к возрастанию отрицательного давления в плевральной полости и к увеличению объема легких, что сопровождается снижением в них давления, оно становится ниже атмосферного и воздух начинает поступать в легкие - происходит вдох.
Между висцеральным и париетальным листками плевры находится щелевидное пространство, которое называется плевральной полостью. Давление в плевральной полости всегда ниже атмосферного, его называют отрицательным давлением. Величина отрицательного давления в плевральной полости равна: к концу максимального выдоха - 1-2 мм рт. ст., к концу спокойного выдоха - 2-3 мм рт. ст., к концу спокойного вдоха -5-7 мм рт. ст., к концу максимального вдоха - 15-20 мм рт. ст.
Отрицательное давление в плевральной полости обусловлено так называемой эластической тягой легких - силой, с которой легкие постоянно стремятся уменьшить свой объем. Эластическая тяга легких обусловлена двумя причинами:
Наличием в стенке альвеол большого количества эластических волокон;
Поверхностным натяжением пленки жидкости, которой покрыта внутренняя поверхность стенок альвеол.
Вещество, покрывающее внутреннюю поверхность альвеол называется сурфактантом. Сурфактант имеет низкое поверхностное натяжение и стабилизирует состояние альвеол, а именно, при вдохе он предохраняет альвеолы от перерастяжения (молекулы сурфактанта расположены далеко друг от друга, что сопровождается повышением величины поверхностного натяжения), а при выдохе - от спадения (молекулы сурфактанта расположены близко друг к другу, что сопровождается снижением величины поверхностного натяжения).
Значение отрицательного давления в плевральной полости в акте вдоха проявляется при поступлении воздуха в плевральную полость, т. е. пневмотораксе. Если в плевральную полость поступает небольшое количество воздуха, легкие частично спадаются, но вентиляция их продолжается. Такое состояние называется закрытым пневмотораксом. Через некоторое время воздух из плевральной полости всасывается и легкие расправляются.
При нарушении герметичности плевральной полости, например, при проникающих ранениях грудной клетки или при разрыве ткани легкого в результате его поражения каким-либо заболеванием, плевральная полость сообщается с атмосферой и давление в ней становится равным атмосферному, легкие спадаются полностью, их вентиляция прекращается. Такой пневмоторакс называется открытым. Открытый двусторонний пневмоторакс несовместим с жизнью.
Частичный искусственный закрытый пневмоторакс (введение в плевральную полость с помощью иглы некоторого количества воздуха) применяется с лечебной целью, например, при туберкулезе частичное спадение пораженного легкого способствует заживлению патологических полостей (каверн).
При глубоком дыхании в акте вдоха участвуют ряд вспомогательных дыхательных мышц, к которым относятся: мышцы шеи, груди, спины. Сокращение этих мышц вызывает перемещение ребер, что оказывает содействие инспираторным мышцам.
При спокойном дыхании вдох осуществляется активно, а выдох пассивно. Силы, обеспечивающие спокойный выдох:
Сила тяжести грудной клетки;
Эластическая тяга легких;
Давление органов брюшной полости;
Эластическая тяга перекрученных во время вдоха реберных хрящей.
В активном выдохе принимают участие внутренние межреберные мышцы, задняя нижняя зубчатая мышца, мышцы живота.
Легочные объемы
Для оценки вентиляционной функции легких, состояния дыхательных путей применяются методы исследования: пневмография, спирометрия, спирография. С помощью спирографа можно определить и записать величины легочных объемов воздуха, проходящих через воздухоносные пути человека.
При спокойном дыхании человек вдыхает и выдыхает около 500 мл воздуха. Этот объем воздуха называется дыхательным объемом . После спокойного вдоха человек может еще максимально вдохнуть некоторое количество воздуха - это резервный объем вдоха , он равен 1800-2000 мл. После спокойного выдоха можно еще максимально выдохнуть некоторое количество воздуха - это резервный объем выдоха , он равен 1300-1500 мл.
Количество воздуха, которое человек может максимально выдохнуть после самого глубокого вдоха называется жизненной емкостью легких (ЖЕЛ). Она складывается из дыхательного объема, резервного объема вдоха и резервного объема выдоха и равна в среднем 3500-4000 мл. Величина ЖЕЛ может изменяться в значительных пределах и зависит от возрастных особенностей организма, степени тренированности человека, наличия сердечно-легочной патологии.
После максимально глубокого выдоха в легких остается некоторое количество воздуха - это остаточный объем , он равен 1300 мл.
Объем воздуха, который находится в легких к концу спокойного выдоха называется функциональной остаточной емкостью , или альвеолярным воздухом. Он состоит из резервного объема выдоха и остаточного объема.
Максимальное количество воздуха, которое может находиться в легких после глубокого вдоха называется общей емкостью легких , она равна сумме остаточного объема и ЖЕЛ.
Воздух находится не только в альвеолах, но и в воздухоносных путях - полости носа, носоглотки, трахеи, бронхов. Воздух, находящийся в воздухоносных путях не участвует в газообмене, поэтому просвет воздухоносных путей называется мертвым пространством. Объем анатомического мертвого пространства около 150 мл.
Хотя в воздухоносных путях не происходит газообмена они необходимы для нормального дыхания, так как в них происходит увлажнение, согревание, очищение от пыли и микроорганизмов вдыхаемого воздуха. При раздражении пылевыми частицами и накопившейся слизью рецепторов носоглотки, гортани и трахеи возникает кашель, а при раздражении рецепторов полости носа - чихание. Кашель и чихание являются защитными дыхательными рефлексами.
Вентиляция легких. Вентиляция легких определяется объемом воздуха, вдыхаемого или выдыхаемого в единицу времени. Количественной характеристикой легочной вентиляции является минутный объем дыхания (МОД) - объем воздуха, проходящего через легкие за одну минуту. В состоянии покоя МОД равен 6-9 л. При физической нагрузке его величина резко возрастает и составляет 25-30 л.
Так как газообмен между воздухом и кровью осуществляется в альвеолах, то важна не общая вентиляция легких, а вентиляция альвеол. Альвеолярная вентиляция меньше вентиляции легких на величину мертвого пространства. Если из величины дыхательного объема вычесть объем мертвого пространства, то получится объем воздуха, содержащегося в альвеолах, а если эту величину умножить на частоту дыхания, получим альвеолярную вентиляцию. Следовательно, эффективность альвеолярной вентиляции выше при более глубоком и редком дыхании, чем при частом и поверхностном.
Состав вдыхаемого, выдыхаемого и альвеолярного воздуха. Атмосферный воздух, которым дышит человек, имеет относительно постоянный состав. В выдыхаемом воздухе меньше кислорода и больше углекислого газа, в альвеолярном воздухе еще меньше кислорода и больше углекислого газа.
Вдыхаемый воздух содержит 20,93% кислорода и 0,03% углекислого газа, выдыхаемый воздух - кислорода 16%, углекислого газа 4,5% и в альвеолярном воздухе содержится 14% кислорода и 5,5% углекислого газа. В выдыхаемом воздухе углекислого газа содержится меньше, чем в альвеолярном. Это связано с тем, что к выдыхаемому воздуху примешивается воздух мертвого пространства с низким содержанием углекислого газа и его концентрация уменьшается.
В дыхательных движениях участвуют 4 анатомо-функциональных образования: воздухоносные дыхательные пути, которые по своим свойствам являются слегка растяжимыми; эластическая и растяжимая легочная ткань; ребра; диафрагма, а также инспираторные и экспираторные мышцы.
Воздухоносный путь - это пространство, которое обеспечивает доставку атмосферного воздуха в альвеолы. Он начинается с отверстий носа и рта, включает полость рта, носоглотку, гортань, трахею, бронхи и бронхиолы до 16-й генерации включительно (они не имеют альвеол). Эти структуры не участвуют в газообмене и составляют анатомическое мертвое пространство; его объем составляет около 150 мл. Бронхиолы 17-19-й генераций образуют переходную зону, их газовая смесь приближается к альвеолярной, а бронхиолы 20-23-й генераций несут на себе основное количество альвеол, в которых происходит газообмен - они образуют дыхательную зону. Хотя в воздухоносных путях нет газообмена, они необходимы для нормального дыхания, так как в них происходит увлажнение, согревание, очищение от пыли и микроорганизмов вдыхаемого воздуха. При раздражении пылевыми частицами и накопившейся слизью рецепторов носоглотки, гортани и трахеи возникает рефлекторный акт кашля, а при раздражении рецепторов полости носа - чиханье. Кашель и чиханье выполняют защитные функции.
Акт вдоха (инспирация) - активный процесс, так как он осуществляется благодаря сокращениям дыхательных мышц, обеспечивающих расширение грудной полости. Мышцы, сокращение которых приводит к увеличению объема грудной полости, называются инспираторными, а мышцы, сокращение которых приводит к уменьшению объема трудной полости, - экспираторными. Основной инспираторной мышцей является мышца диафрагмы; при сокращении ее купол уплощается при спокойном дыхании на 1,5-2 см, при глубоком - до 10 см. При этом происходит смещение внутренних органов вниз, что приводит к увеличению объема грудной клетки в вертикальном направлении. В осуществлении спокойного вдоха, кроме диафрагмы, также принимают участие наружные межреберные и межхрящевые мышцы, сокращение которых приводит к увеличению объема грудной клетки в сагиттальном и фронтальном направлениях. При форсированном вдохе вспомогательную функцию выполняют мышцы, прикрепленные к костям плечевого пояса, черепу или позвоночнику и способные поднимать ребра: грудино-ключично-сосцевидная, трапециевидная, обе грудные мышцы, мышца, поднимающая лопатку, лестничная, передняя зубчатая.
Большое значение в процессе дыхания имеет отрицательное давление в плевральной полости (плевральной щели). Каждое легкое покрыто серозной оболочкой - плеврой, состоящей из висцерального и париетального листков.
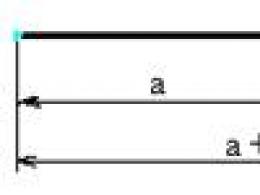
В паузе между вдохом и выдохом атмосферное давление (Ратм.), действующее на стенку альвеол изнутри, уравновешено суммой внутриплеврального давления (Р пл.) и эластической тягой легких (Р эл.), возникающей при растяжении легочной ткани: Р атм = Р пл + Р эл. При увеличении объема грудной клетки в результате сокращения инспираторных мышц париетальный листок следует за грудной клеткой. Это приводит к уменьшению давления в плевральной щели. В результате атмосферное давление растягивает ткань легких: Р атм > Р пл +Р эл. Поэтому висцеральный листок, а вместе с ним и легкие следуют за париетальным листком. Воздух начинает поступать в легкие, происходит вдох. Легкие растягиваются до тех пор, пока атмосферное давление вновь не будет уравновешено суммой плеврального давления и эластической тяги.
Давление в плевральной полости всегда отрицательное (ниже атмосферного). Величина отрицательного давления в плевральной полости неодинакова в разные фазы дыхания: к концу максимального выдоха она равна 1-2 мм рт. ст., к концу спокойного выдоха - 2-3 мм рт. ст., к концу спокойного вдоха - 5-7 мм рт. ст., к концу максимального вдоха - 15-20 мм рт. ст.
Отрицательное давление в плевральной полости обусловлено так называемой эластической тягой легких - силой, с которой легкие постоянно стремятся уменьшить свой объем. Эластическая тяга обусловлена двумя причинами: наличием в стенке альвеол большого количества эластических волокон; поверхностным натяжением пленки жидкости, которой покрыта внутренняя поверхность стенок альвеол. (Сурфактант).
При вдохе он предохраняет альвеолы от перерастяжения, так как молекулы сурфактанта расположены далеко друг от друга, что сопровождается повышением величины поверхностного натяжения. При выдохе он предохраняет альвеолы от спадения, так как молекулы сурфактанта расположены близко друг к другу, что сопровождается снижением величины поверхностного натяжения.
В отличие от спокойного вдоха спокойный выдох - пассивный процесс: он происходит без участия экспираторных мышц на фоне расслабления инспираторной мускулатуры за счет энергии, которая накопилась во время вдоха. Для осуществления спокойного выдоха обычно достаточно эластических свойств легких и массы переместившихся во время вдоха тканей.
Спокойный выдох обеспечивается следующими силами: массой грудной клетки, возвращающейся к исходному состоянию под действием силы тяжести; эластической тягой легких; давлением органов брюшной полости; эластической тягой перекрученных во время вдоха реберных хрящей.
При этом Р атм < Р пл + Р эл., что приводит к уменьшению объема легких и изгнанию части воздуха в атмосферу.В форсированном выдохе принимают участие внутренние межреберные мышцы, задняя нижняя зубчатая мышца, мышцы живота.
Билет № 4
Функциональная система - по П.К.Анохину - комплекс избирательно извлеченных компонентов организма, взаимодействия и взаимоотношения которых ориентированы на получение фокусированного полезного результата.
Функциональная система: -является единица интегративной деятельности целого организма; -отличается от частных механизмов реализации поведенческих актов; -осуществляет избирательное вовлечение структур и процессов в выполнение конкретного акта поведения или функции организма; - имеет разветвленный морфофизиологический аппарат, обеспечивающий поддержание гомеостаза и саморегуляцию.
Различают функциональные системы первого и второго типов.
Выделяют два типа функциональных систем. 1. Функциональные системы первого типа обеспечивают постоянство определенных констант внутренней среды за счет системы саморегуляции, звенья которой не выходят за пределы самого организма. Примером может служить функциональная система поддержания постоянства кровяного давления, температуры тела и т.п. Такая система с помощью разнообразных механизмов автоматически компенсирует возникающие сдвиги во внутренней среде. 2. Функциональные системы второго типа используют внешнее звено саморегуляции. Они обеспечивают приспособительный эффект благодаря выходу за пределы организма через связь с внешним миром, через изменения поведения. Именно функциональные системы второго типа лежат в основе различных поведенческих актов, различных типов поведения.
2. Микроциркуля́ция ) - транспорт биологических жидкостей на тканевом уровне. Это понятие включает в себя капиллярное кровообращение (движение крови по микрососудам капиллярного типа)Капилляры представляют собой тончайшие сосуды, диаметром 5-7 мкм, длиной 0,5-1,1 мм. Эти сосуды пролегают в межклеточных пространствах, тесно соприкасаясь с клетками органов и тканей организма. Суммарная длина всех капилляров тела человека составляет около 100 000 км, т. е. нить, которой можно было бы 3 раза опоясать земной шар по экватору. Физиологическое значение капилляров состоит в том, что через их стенки осуществляется обмен веществмежду кровью и тканями. Стенки капилляров образованы только одним слоем клеток эндотелия, снаружи которого находится тонкая соединительнотканная базальная мембрана.Скорость кровотока в капиллярах невелика и составляет 0,5- 1 мм/с. Таким образом, каждая частица крови находится в капилляре примерно 1 с. Небольшая толщина слоя крови (7-8 мкм) и тесный контакт его с клетками органов и тканей, а также непрерывная смена крови в капиллярах обеспечивают возможность обмена веществ между кровью и тканевой (межклеточной) жидкостью.В тканях, отличающихся интенсивным обменом веществ, число капилляров на 1 мм2 поперечного сечения больше, чем в тканях, в которых обмен веществ менее интенсивный. Так, в сердце на 1 мм2 сечения в 2 раза больше капилляров, чем в скелетноймышце. В сером веществе мозга, где много клеточных элементов, капиллярная сеть значительно более густая, чем в белом.
Различают два вида функционирующих капилляров. Одни из них образуют кратчайший путь между артериолами и венулами (магистральные капилляры). Другие представляют собой боковые ответвления от первых: они отходят от артериального конца магистральных капилляров и впадают в их венозный конец. Эти боковые ответвления образуют капиллярные сети. Объемная и линейная скорость кровотока в магистральных капиллярах больше, чем в боковых ответвлениях. Магистральные капилляры играют важную роль в распределении крови в капиллярных сетях и в других феноменах микроциркуляции.Кровь течет лишь в «дежурных» капиллярах. Часть капилляров выключена из кровообращения. В период интенсивной деятельности органов (например, при сокращении мышц или секреторной активности желез), когда обмен веществ в них усиливается, количество функционирующих капилляров значительно возрастает.Регулирование капиллярного кровообращения нервной системой, влияние на него физиологически активных веществ - гормонов и метаболитов - осуществляются при воздействии их на артерии и артериолы. Сужение или расширение артерий и артериол изменяет как количество функционирующих капилляров, распределение крови в ветвящейся капиллярной сети, так и состав крови, протекающей по капиллярам, т. е. соотношение эритроцитов и плазмы. При этом общий кровоток через метартериолы и капилляры определяется сокращением гладких мышечных клеток артериол, а степень сокращения прекапиллярных сфинктеров (гладких мышечных клеток, расположенных у устья капилляра при его отхождении от метаартериол) определяет, какая часть крови пройдет через истинные капилляры.
В некоторых участках тела, например в коже, легких и почках, имеются непосредственные соединения артериол и венул - артериовенозные анастомозы. Это наиболее короткий путь между артериолами и венулами. В обычных условиях анастомозы закрыты и кровь проходит через капиллярную сеть. Если анастомозы открываются, то часть крови может поступать в вены, минуя капилляры.Артериовенозные анастомозы играют роль шунтов, регулирующих капиллярное кровообращение. Примером этого является изменение капиллярного кровообращения в коже при повышении (свыше 35°С) или понижении (ниже 15°С) температуры окружающей среды. Анастомозы в коже открываются и устанавливается ток крови из артериол непосредственно в вены, что играет большую роль в процессах терморегуляции.Структурной и функциональной единицей кровотока в мелких сосудах является сосудистый модуль - относительно обособленный в гемодинамическом отношении комплекс микрососудов, снабжающий кровью определенную клеточную популяцию органа. При этом имеет место специфичность васкуляризации тканей различных органов, что проявляется в особенностях ветвления микрососудов, плотности капилляризации тканей и др. Наличие модулей позволяет регулировать локальный кровоток в отдельных микроучастках тканей. Оно объединяет механизмы кровотока в мелких сосудах и теснейшим образом связанный с кровотоком обмен жидкостью и растворенными в ней газами и веществами между сосудами и тканевой жидкостью.Специального рассмотрения заслуживают процессы обмена между кровью и тканевой жидкостью. Через сосудистую систему за сутки проходит 8000-9000 л крови. Через стенку капилляров профильтровывается около 20 л жидкости и 18 л реабсорбируется в кровь. По лимфатическим сосудам оттекает около 2 л жидкости. Закономерности, обусловливающие обмен жидкости между капиллярами и тканевыми пространствами, были описаны Стерлингом. Гидростатическое давление крови в капиллярах (Ргк) является основной силой, направленной на перемещение жидкости из капилляров в ткани.
3. Почки выполняют ряд гомеостатических функций в организме человека и высших животных. К функциям почек относятся следующие: 1) участие в регуляции объема крови и внеклеточной жидкости (волюморегуляция); 2) регуляция концентрации осмотически активных веществ в крови и других жидкостях тела (осморегуляция); 3) регуляция ионного состава сыворотки крови и ионного баланса организма {ионная регуляция); 4) участие в регуляции кислотно-основного состояния {стабилизация рН крови)", 5) участие в регуляции артериального давления, эритропоэза, свертывания крови, модуляции действия гормонов благодаря образованию и выделению в кровь биологически активных веществ (инкреторная функция); 6) участие в обмене белков, липидов и углеводов (метаболическая функция); 7) выделение из организма конечных продуктов азотистого обмена и чужеродных веществ, избытка органических веществ (глюкоза, аминокислоты и др.), поступивших с пищей или образовавшихся в процессе метаболизма (экскреторная функция). Таким образом, роль почки в организме не ограничивается только выделением конечных продуктов обмена и избытка неорганических и органических веществ. Почка является гомеостатическим органом, участвующим в поддержании постоянства основных физико-химических констант жидкостей внутренней среды, в циркуляторном гомеостазе, стабилизации показателей обмена различных органических веществ.При изучении работы почки следует разграничить два понятия - функции почки и процессы, их обеспечивающие. К последним относятся ультрафильтрация жидкости в клубочках, реабсорбция и секреция веществ в канальцах, синтез новых соединений, в том числе и биологически активных веществ (рис. 12.1).В литературе при описании деятельности почки используют термин «секреция», который имеет ряд значений. В одних случаях этот термин означает перенос вещества клетками нефрона из крови в просвет канальца в неизмененном виде, что обусловливает экскрецию этого вещества почкой. В других случаях термин «секреция» означает синтез и секрецию клетками в почке биологически активных веществ (например, ренина, простагландинов) и их поступление в русло крови. Наконец, процесс синтеза в клетках канальцев веществ, которые поступают в просвет канальца и экскретируются с мочой, также обозначают термином «секреция».
Строение нефрона. В каждой почке у человека содержится около 1 млн функциональных единиц - нефронов, в которых происходит образование мочи (рис. 12.2). Каждый нефрон начинается почечным, или мальпигиевым, тельцем - двустенной капсулой клубочка (капсула Шумлянского-Боумена), внутри которой находится клубочек капилляров. Внутренняя поверхность капсулы выстлана эпителиальными клетками; образующаяся полость между висцеральным и париетальным листками капсулы переходит в просвет проксимального извитого канальца. Особенностью клеток этого канальца является наличие щеточной каемки - большого количества микроворсинок, обращенных в просвет канальца. Следующий отдел нефрона - тонкая нисходящая часть петли нефрона (петли Генле). Ее стенка образована низкими, плоскими эпителиальными клетками. Нисходящая часть петли может опускаться глубоко в мозговое вещество, где каналец изгибается на 180°, и поворачивает в сторону коркового вещества почки, образуя восходящую часть петли нефрона. Она может включать тонкую и всегда имеет толстую восходящую часть, которая поднимается до уровня клубочка своего же нефрона, где начинается дистальный извитой каналец. Этот отдел канальца обязательно прикасается к клубочку между приносящей и выносящей артериолами в области плотного пятна (см. рис. 12.2). Клетки толстого восходящего отдела петли Генле и дистального извитого канальца лишены щеточной каемки, в них много митохондрий и увеличена поверхность базальной плазматической мембраны за счет складчатости. Конечный отдел нефрона - короткий связующий каналец, впадает в собирательную трубку1. Начинаясь в корковом веществе почки, собирательные трубки проходят через мозговое вещество и открываются в полость почечной лоханки. Диаметр капсулы клубочка около 0,2 мм, общая длина канальцев одного нефрона достигает 35-50 мм. Исходя из особенностей структуры и функции почечных канальцев, различают следующие сегменты нефрона: 1) проксимальный, в состав которого входят извитая и прямая части проксимального канальца; 2) тонкий отдел петли нефрона, включающий нисходящую и тонкую восходящую части петли; 3) дистальный сегмент, образованный толстым восходящим отделом петли нефрона, дистальным извитым канальцем и связующим отделом. Канальцы нефрона соединены с собирательными трубками: в процессе эмбриогенеза они развиваются самостоятельно, но в сформировавшейся почке собирательные трубки функционально близки дистальному сегменту нефрона.
В почке функционирует несколько типов нефронов: суперфициальные (поверхностные), интракортикальные и юкстамедуллярные (см. рис. 12.2). Различие между ними заключается в локализации в почке, величине клубочков (юкстамедуллярные крупнее суперфициальных), глубине расположения клубочков и проксимальных канальцев в корковом веществе почки (клубочки юкстамедуллярных нефронов лежат у границы коркового и мозгового вещества) и в длине отдельных участков нефрона, особенно петель нефрона. Суперфициальные нефроны имеют короткие петли, юкстамедуллярные, напротив, длинные, спускающиеся во внутреннее мозговое вещество почки. Характерна строгая зональность распределения канальцев внутри почки Большое функциональное значение имеет зона ночки, в которой расположен каналец, независимо от того, находится ли он в корковом или мозговом веществе. В корковом веществе находятся почечные клубочки, проксимальные и дистальные отделы канальцев, связующие отделы. В наружной полоске наружного мозгового вещества находятся нисходящие и толстые восходящие отделы петель нефронов, собирательные трубки; во внутреннем мозговом веществе располагаются тонкие отделы петель нефронов и собирательные трубки. Расположение каждой из частей нефрона в почке чрезвычайно важно и определяет форму участия тех или иных нефронов в деятельности почки, в частности в осмотическом концентрировании мочи. Кровоснабжение почки. В обычных условиях через обе почки, масса которых составляет лишь около 0,43 % от массы тела здорового человека, проходит от 1/5 до 1/44 крови, поступающей из сердца в аорту. Кровоток по корковому веществу почки достигает 4-5 мл/мин на 1 г ткани; это наиболее высокий уровень органного кровотока. Особенность почечного кровотока состоит в том, что в условиях изменения системного артериального давления в широких пределах (от 90 до 190 мм рт. ст.) он остается постоянным. Это обусловлено специальной системой саморегуляции кровообращения в почке.
Короткие почечные артерии отходят от брюшного отдела аорты, разветвляются в почке на все более мелкие сосуды, и одна приносящая (афферентная) артериола входит в клубочек. Здесь она распадается на капиллярные петли, которые, сливаясь, образуют выносящую (эфферентную) артериолу, по которой кровь оттекает от клубочка. Диаметр эфферентной артериолы уже, чем афферентной. Вскоре после отхождения от клубочка эфферентная артериола вновь распадается на капилляры, образуя густую сеть вокруг проксимальных и дистальных извитых канальцев. Таким образом, большая часть крови в почке дважды проходит через капилляры - вначале в клубочке, затем у канальцев. Отличие кровоснабжения юкстамедуллярного нефрона заключается в том, что эфферентная артериола не распадается на околоканальцевую капиллярную сеть, а образует прямые сосуды, спускающиеся в мозговое вещество почки. Эти сосуды обеспечивают кровоснабжение мозгового вещества почки; кровь из околоканальцевых капилляров и прямых сосудов оттекает в венозную систему и по почечной вене поступает в нижнюю полую вену Юкстагломерулярный аппарат (рис. 12.3). Морфологически образует подобие треугольника, две стороны которого представлены подходящими к клубочку афферентной и эфферентной артериолами, а основание - клетками плотного пятна (mucula densa) дистального канальца. Внутренняя поверхность афферентной артериолы выстлана эндотелием, а мышечный слой вблизи клубочка замещен крупными эпителиальными клетками, содержащими секреторные гранулы. Клетки плотного пятна тесно соприкасаются с юкстагломерулярным веществом, состоящим из ячеистой сети с мелкими клетками и переходящим в клубочек, где расположена мезангиальная ткань. Юкстагломерулярный аппарат участвует в секреции ренина и ряда других биологически активных веществ.
Билет № 5 1. Рефлекс (от лат. reflexus отражённый) стереотипная реакция живого организма на определенное воздействие, проходящая с участием нервной системы. Рефлексы существуют у многоклеточных живых организмов, обладающих нервной системой. Классификация:
По типу образования: условные и безусловные.
По видам рецепторов: экстероцептивные (кожные, зрительные, слуховые, обонятельные), интероцептивные (с рецепторов внутренних органов) и проприоцептивные (с рецепторов мышц, сухожилий, суставов)
По эффекторам: соматические, или двигательные, (рефлексы скелетных мышц), например флексорные, экстензорные, локомоторные, статокинетические и др.; вегетативные внутренних органов - пищеварительные, сердечно-сосудистые, выделительные, секреторные и др.
По биологической значимости: оборонительные, или защитные, пищеварительные, половые, ориентировочные.
По степени сложности нейронной организации рефлекторных дуг различают моносинаптические, дуги которых состоят из афферентного и эфферентного нейронов (например, коленный), и полисинаптические, дуги которых содержат также 1 или несколько промежуточных нейронов и имеют 2 или несколько синаптических переключений (например, флексорный).
По характеру влияний на деятельность эффектора: возбудительные - вызывающими и усиливающими (облегчающими) его деятельность, тормозные - ослабляющими и подавляющими её (например, рефлекторное учащение сердечного ритма симпатическим нервом и урежение его или остановка сердца - блуждающим).
По анатомическому расположению центральной части рефлекторных дуг различают спинальные рефлексы и рефлексы головного мозга. В осуществлении спинальных рефлексов участвуют нейроны, расположенные в спинном мозге. Пример простейшего спинального рефлекса - отдергивание руки от острой булавки. Рефлексы головного мозга осуществляются при участии нейронов головного мозга. Среди них различают бульбарные, осуществляемые при участии нейронов продолговатого мозга; мезэнцефальные - с участием нейронов среднего мозга; кортикальные - с участием нейронов коры больших полушарий головного мозга.
Рефлекторная дуга (нервная дуга) - путь, проходимый нервными импульсами при осуществлении рефлекса.
Рефлекторная дуга состоит из: рецептора - нервное звено, воспринимающее раздражение; афферентного звена - центростремительное нервное волокно - отростки рецепторных нейронов, осуществляющие передачу импульсов от чувствительных нервных окончаний в центральную нервную систему; центрального звена - нервный центр (необязательный элемент, например для аксон-рефлекса); эфферентного звена - осуществляют передачу от нервного центра к эффектору. эффектора - исполнительный орган, деятельность которого изменяется в результате рефлекса.
Различают: - моносинаптические, двухнейронные рефлекторные дуги; полисинаптические рефлекторные дуги (включают три и более нейронов).
Рефлекторное кольцо - совокупность структур нервной системы, участвующих в осуществлении рефлекса и передаче информации о характере и силе рефлекторного действия в центральной нервной системе. Рефлекторное кольцо включает в себя: рефлекторную дугу; обратную афферентацию от эффекторного органа в центральную нервную систему.
Смысл обратной афферентации состоит в том, что в любом физиологическом процессе или в поведенческом акте животного, который направлен на получение какого-то приспособительного эффекта, обратная афферентация информирует о результатах совершенного действия, давая возможность организму оценить степень успеха выполняемого им действия, т.е. через звено обратной афферентации осуществляется постоянная оценка реально полученного результата с тем, который был «запрограммирован» в акцепторе результата действия.
2) Цикл сердечной деятельности состоит из систолы и диастолы. Систола – сокращение, которое длится 0,1–0,16 с в предсердии и 0,3–0,36 с в желудочке. Систола предсердий слабее, чем систола желудочков. Диастола – расслабление, у предсердий занимает 0,7–0,76 с, у желудочков – 0,47-0,56 с. Продолжительность сердечного цикла составляет 0,8–0,86 с и зависит от частоты сокращений. Время, в течение которого предсердия и желудочки находятся в состоянии покоя, называется общей паузой в деятельности сердца. Она длится примерно 0,4 с. В течение этого времени сердце отдыхает, а его камеры частично наполняются кровью. Систола и диастола – сложные фазы и состоят из нескольких периодов. В систоле различают два периода – напряжения и изгнания крови, включающие в себя:
1) фазу асинхронного сокращения – 0,05 с;
2) фазу изометрического сокращения – 0,03 с;
3) фазу быстрого изгнания крови – 0,12 с;
4) фазу медленного изгнания крови – 0,13 с.
Диастола продолжается около 0,47 с и состоит из трех периодов:
1) протодиастолического – 0,04 с;
2) изометрического – 0,08 с;
3) периода наполнения, в котором выделяют фазу быстрого изгнания крови – 0,08 с, фазу медленного изгнания крови – 0,17 с, время пресистолы – наполнение желудочков кровью – 0,1 с.
В общем можем сказать, что основными параметрами, характеризующими системную гемодинамику, являются: системное артериальное давление, общее периферическое сопротивление сосудов, сердечный выброс, работа сердца, венозный возврат крови к сердцу, центральное венозное давление, объем циркулирующей крови.
3) Неорганические вещества плазмы крови:
Катионы: Na+, К+, Са2+, Mg2+, Fe3+, Cu2+; - анионы Cl-, PO43-, HCO3-, I-. Значение: обеспечение осмотического давления крови (на 60 % - NaCl). В норме давление осмотическое крови равно 7,7-8,1 атм.; обеспечение pH крови равное 7,36-7,4; обеспечение определенного уровня чувствительности клеток, участвующих в формировании мембранного потенциала.
Осмотическое давление - избыточное гидростатическое давление на раствор, отделённый от чистого растворителя полупроницаемой мембраной, при котором прекращается диффузия растворителя через мембрану. Это давление стремится уравнять концентрации обоих растворов вследствие встречной диффузии молекул растворённого вещества и растворителя. Раствор, имеющий более высокое осмотическое давление по сравнению с другим раствором, называется гипертоническим, имеющий более низкое - гипотоническим. Значение осм. давл крови- 7,6 - 8,1 атм. Оно создается в основном солями, находящимися в диссоциированном состоянии. Осмотическое давление имеет существенное значение в поддержании концентрации различных веществ, растворенных в жидкостях организма, и определяет распределение воды между кровью, клетками и тканями.
Изотонический раствор - это раствор, осмотическое давление которого равно осмотическому давлению крови (например, 0,85% раствор NaCl). Эритроциты, помещенные в такой раствор, не изменяются, так как осмотическое давление в них и в растворе одинаково. Данный раствор получил название - физиологический. Гипотонический раствор - это раствор, осмотическое давление которого ниже осмотического давления крови (например 0,3% раствор NaCI). Эритроциты, помещенные в такой раствор, набухают и лопаются (гемолизируются) в результате перехода воды в клетку, так как осмотическое давление в эритроците выше, чем в растворе. Гипертонический раствор - это раствор, осмотическое давление которого выше осмотического давления крови (например, 2% раствор NaCI). Эритроциты, помещенные в такой раствор, сморщиваются в результате выхода воды из клетки, так как осмотическое давление в эритроцитах ниже, чем в растворе.
Функциональная система, обеспечивающая постоянство осмотического давления крови.
1) Спинной мозг (medulla spinalis) - часть центральной нервной системы, расположенная в позвоночном канале.
Спинной мозг выполняет рефлекторную и проводниковую функции. Первая обеспечивается его нервными центрами, вторая проводящими путями.
Он имеет сегментарное строение. Причем деление на сегменты является функциональным. Каждый сегмент образует передние и задние корешки. Задние являются чувствительными, т.е. афферентными, передние двигательными, эфферентными. Корешки каждого сегмента иннервируют 3 метамера тела, но в результате перекрывания каждый метамер иннервируется тремя сегментами. Поэтому при поражении передних корешков одного сегмента, двигательная активность соответствующего метамера лишь ослабляется.
Морфологически тела нейронов спинного мозга образуют его серое вещество. Функционально все его нейроны делятся на мотонейроны, вставочные, нейроны симпатического и парасимпатического отделов вегетативной нервной системы.
Мотонейроны , в зависимости от функционального значения, разделяются на альфа – и гамма-мотонейроны. К a-мотонейронам идут волокна афферентных путей, которые начинаются от интрафузальных, т.е. рецепторных мышечных клеток. Тела a-мотонейронов расположены в передних рогах спинного мозга, а их аксоны иннервируют скелетные мышцы. Гамма-мотонейроны регулируют напряжение мышечных веретен т.е. интрафузальных волокон. Таким образом они участвуют в регуляции сокращений скелетных мышц. Поэтому при перерезке передних корешков мышечный тонус исчезает
Интернейроны обеспечивают связь между центрами спинного мозга и вышележащих отделов ЦНС.
Нейроны симпатического отдела вегетативной нервной системы находятся в боковых рогах грудных сегментов, а парасимпатического в крестцовом отделе.
Проводниковая функция состоит в обеспечении связи периферических рецепторов, центров спинного мозга с вышележащими отделами ЦНС, а также его нервных центров между собой. Она осуществляется проводящими путями. Все пути спинного мозга делятся на собственные или проприоспинальные, восходящие и нисходящие. Проприоспинальные пути связывают между собой нервные центры разных сегментов спинного мозга. Их функция заключается в координации тонуса мышц, движений различных метамеров туловища. К восходящим путям относятся несколько трактов. Пучки Голля и Бурдаха проводят нервные импульсы от проприорецепторов мышц и сухожилий к соответствующим ядрам продолговатого мозга, а затем таламусу и соматосенсорным зонам коры. Благодаря этим путям производится оценка и коррекция позы туловища. Пучки Говерса и Флексига передают возбуждение от проприорецепторов, механорецепторов кожи к мозжечку. За счет этого обеспечивается восприятие и бессознательная координация позы. Спиноталамические тракты проводят сигналы от болевых, температурных, тактильных рецепторов кожи к таламусу, а затем соматосенсорные зоны коры. Они обеспечивают восприятие соответствующих сигналов и формирование чувствительности. Нисходящие пути также образованы несколькими трактами. Кортикоспинальные пути идут от пирамидных и экстрапирамидных нейронов коры к a-мотонейронам спинного мозга. За счет них осуществляется регуляция произвольных движений. Руброспинальный путь проводит сигналы от красного ядра среднего мозга к a-мотонейронам мышц сгибателей. Вестибулоспинальный путь передает сигналы от вестибулярных ядер продолговатого мозга, в первую очередь ядра Дейтерса, к a-мотонейронам мышц разгибателей. За счет этих двух путей регулируется тонус соответствующих мышц при изменениях положения тела.
Рефлекторная функция
Все рефлексы спинного мозга делятся на соматические, т.е. двигательные и вегетативные.
Соматические рефлексы делятся на сухожильные или миотатические и кожные. Сухожильные рефлексы возникают при механическом раздражении мышц и сухожилий. Их небольшое растяжение приводит к возбуждению сухожильных рецепторов и a-мотонейронов спинного мозга. В результате возникает сокращение мышц, в первую очередь разгибателей. К сухожильным рефлексам относятся коленный, ахиллов, локтевой, кистевой и др., возникающие при механическом раздражении соответствующих сухожилий. Например, коленный является простейшим моносинаптическим, так как в его центральной части только один синапс. Кожные рефлексы обусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной (объяснение). Спинальные нервные центры находятся под контролем вышележащих. Поэтому после перерезки между продолговатым и спинным мозгом возникает спинальный шок и тонус всех мышц значительно уменьшится.
Вегетативные рефлексы спинного мозга делятся на симпатические и парасимпатические. Те и другие проявляются реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны спинного мозга образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыведения, дефекации, эрекции, эйякуляции и т.д.
2) Дыхательный центр расположен в медиальной части ретикулярной формации продолговатого мозга. Его верхняя граница находится ниже ядра лицевого нерва, а нижняя выше писчего пера.
Этот центр состоит из инспираторных и экспираторных нейронов . В первых нервные импульсы начинают генерироваться незадолго до вдоха и продолжаются в течение всего вдоха. Несколько ниже расположенные экспираторные нейроны. Они возбуждаются к концу вдоха и находятся в возбужденном состоянии в течение всего выдоха. В инспираторном центре имеется 2 группы нейронов. Это респираторные a – и b-нейроны. Первые возбуждаются при вдохе. Одновременно к b-респираторным нейронам поступают импульсы от экспираторных. Они активируются одновременно с a-респираторными нейронами и обеспечивают их торможение в конце вдоха. Благодаря этим связям нейронов дыхательного центра они находятся в реципрокных отношениях (т.е. при возбуждении инспираторных нейронов экспираторные тормозятся и наоборот). Кроме того нейронам бульбарного дыхательного центра свойственно явление автоматии. Это их способность даже в отсутствии нервных импульсов от периферических рецепторов генерировать ритмические разряды биопотенциалов.
Благодаря автоматии дыхательного центра происходит самопроизвольная смена фаз дыхания. Автоматия нейронов объясняется ритмическими колебаниями обменных процессов в них, а также воздействием на них углекислого газа. Эфферентные пути от бульбарного дыхательного центра идут к мотонейронам дыхательных межреберных и диафрагмальных мышц. Мотонейроны диафрагмальных мышц находятся в передних рогах 3-4 шейных сегментов спинного мозга, а межреберных в передних рогах грудных сегментов. Вследствие этого перерезка на уровне 1-2 шейных сегментов ведет к прекращению сокращений дыхательных мышц. В передней части варолиева моста также имеются группы нейронов участвующих в регуляции дыхания. Эти нейроны имеют восходящие и нисходящие связи с нейронами бульбарного центра. К ним идут импульсы от его инспираторных нейронов, а от них к экспираторным. За счет этого обеспечивается плавный переход от вдоха к выдоху, а также координация длительности фаз дыхания.
Апноэстический центр угнетения дыхания находится в нижней части моста и обеспечивает достаточную длительность вдоха. Хотя до сих пор неясно, играет ли этот центр роль в регуляции дыхания у человека, доказано его участие в увеличении длительности вдоха.
Пневмотаксический центр , расположенный в парабрахиальном ядре в верхней части моста, обеспечивает прекращение вдоха (т.е. ограничивает вдох). Более выраженная активность этого центра вызывает укорочение вдоха (до 0,5 с) и учащение дыхания до 30-40 вдохов в минуту, а слабая активность - длительный вдох (до 5 с и более) и снижение частоты дыхания до нескольких вдохов в минуту. Пневмотаксический центр участвует в регуляции объема вдоха.
3) Гру́ппа кро́ви - описание индивидуальных антигенных характеристик эритроцитов, определяемое с помощью методов идентификации специфических групп углеводов и белков, включённых в мембраны эритроцитов.
Группы крови системы АВО обозначаются римскими цифрами и дублирующим названием антигена:
I(0) – в эритроцитах нет агглютиногенов, но в плазме содержатся агглютинины a и b.
II(A) – агглютиногены А и агглютинины b.
III(B) – агглютиногены В и агглютинины a.
IV(AB) – в эритроцитах агглютиногены А и В, агглютининов в плазме нет.
Резус крови - это антиген (белок), который находится на поверхности красных кровяных телец (эритроцитов). В отличие от антигенной системы АВО, где к агглютиногенам А и В имеются соответствующие агглютинины, агглютининов к резус-антигену в крови нет. Они вырабатываются в том случае, если резус-положительную кровь (содержащую резус-фактор) перелить реципиенту с резус-отрицательной кровью. При первом переливании резус несовместимой крови никакой трансфузионной реакции не будет. Однако в результате сенсибилизации организма реципиента, через 3-4 недели в его крови появятся резус-агглютинины. Они очень длительное время сохраняются. Поэтому при повторном переливании резус-положительной крови этому реципиенту произойдет агглютинация и гемолиз эритроцитов донорской крови.
Другое отличие этих двух антигенных систем состоит в том, что резус-агглютинины имеют значительно меньшие размеры, чем a и b. Поэтому они могут проникать через плацентарный барьер. В последние недели беременности, во время родов и даже при абортах, эритроциты плода могут попадать в кровяное русло матери. Если плод имеет резус-положительную кровь, а мать резус-отрицательную, то попавшие в ее организм с эритроцитами плода резус-антигены, вызовут образование резус-агглютининов. Титр резус-агглютининов нарастает медленно, поэтому при первой беременности особых осложнений не возникает. Если при повторной беременности плод опять наследует резус-положительную кровь, то поступающие через плаценту резус-агглютинины матери вызовут агглютинацию и гемолиз эритроцитов плода. В легких случаях возникает анемия, гемолитическая желтуха новорожденных. В тяжелых эритробластоз плода и мертворожденность. Это явление называется резус-конфликтом.
В настоящее время известно около 400 антигенных систем крови. Кроме систем АВО и Rh, известны систем MNSs, Р, Келла, Кидда и другие. Учитывая все антигены число их комбинаций составляет около 300 млн. Но так как их антигенные свойства выражены слабо, для переливания крови их роль незначительна.
Переливание крови (гемотрансфузия) - лечебный метод, заключающийся во введении в кровеносное русло больного (реципиента) цельной крови или ее компонентов, заготовленных от донора или от самого реципиента (аутогемотрансфузия), а также крови, излившейся в полости тела при травмах и операциях (реинфузия).
Показания к назначению переливания любой трансфузионной среды, а также ее дозировка и выбор метода трансфузии определяются лечащим врачом на основании клинических и лабораторных данных. Врач обязан независимо от проведенных ранее исследований и имеющихся записей лично провести следующие контрольные исследования:
1) определить групповую принадлежность крови реципиента по системе AB0 и сверить результат с данными истории болезни;
2) определить групповую принадлежность эритроцитов донора и сопоставить результат с данными на этикетке контейнера или бутылки;
3) провести пробы на совместимость в отношении групп крови донора и реципиента по системе AB0 и резус-фактору;
4) провести биологическую пробу.
Билет № 7
1 Типы ВНД - совокупность врожденных (генотип) и приобретенных (фенотип) свойств нервной системы, определяющих характер взаимодействия организма с окружающей средой и находящих свое отражение во всех функциях организма.
Различные комбинации трех основных свойств нервной системы - силы процессов возбуждения и торможения, их уравновешенности и подвижности - позволили И.П. Павлову выделить четыре резко очерченных типа, отличающихся по адаптивным способностям и устойчивости к невротизирующим агентам.
Т.ВНД сильный неуравновешенный - характеризуется сильным раздражительным процессом и отстающим по силе тормозным, поэтому представитель такого типа в трудных ситуациях легко подвержен нарушениям ВНД. Способен тренировать и в значительной степени улучшать недостаточное торможение. В соответствии с учением о темпераментах - это холерический тип.
Т. ВНД уравновешенный инертный - с сильными процессами возбуждения и торможения и с плохой их подвижностью, всегда испытывающий затруднения при переключении с одного вида деятельности на другой. В соответствии с учением о темпераментах - это флегматический тип.
Т.ВНД сильный уравновешенный подвижный - имеет одинаково сильные процессы возбуждения и торможения с хорошей их подвижностью, что обеспечивает высокие адаптивные возможности и устойчивость в условиях трудных жизненных ситуаций. В соответствии с учением о темпераментах - это сангвинический тип.
Т.ВНД слабый - характеризуется слабостью обоих нервных процессов - возбуждения и торможения, плохо приспосабливается к условиям окружающей среды, подвержен невротическим расстройствам. В соответствии с классификацией темпераментов - это меланхолический тип.
Темперамент - устойчивое объединение индивидуальных особенностей личности, связанных с динамическими, а не содержательными аспектами деятельности. Темперамент составляет основу развития характера; вообще, с физиологической точки зрения темперамент - тип высшей нервной деятельности человека.
Темперамент по Гиппократу:
Преобладание жёлтой жёлчи делает человека импульсивным, «горячим» - холериком.
Преобладание лимфы делает человека спокойным и медлительным - флегматиком.
Преобладание крови делает человека подвижным и весёлым - сангвиником.
Преобладание чёрной жёлчи делает человека грустным и боязливым - меланхоликом.
Конституция - это совокупность функциональных и морфологических особенностей организма, сложившихся на основе наследственных и приобретенных свойств, которые определяют своеобразие реакции организма на внешние и внутренние раздражители.
Типология человека Кречмера:
Астеник - отличается слабым ростом «в толщину» при большем росте «в длину»; худой, тонкий, с бедной соками и кровью кожей, узкими плечами, длинной и плоской грудной клеткой. Имеет хрупкое телосложение, высокий рост, вытянутое лицо, длинный тонкий нос. Нижние конечности длинные и худые. Астенические женщины напоминают астеников-мужчин, но они не только худощавы, но и малорослы. Бросается в глаза их преждевременное старение.
Пикник - среднего или малого роста, с богатой жировой тканью, расплывшимся туловищем, круглой головой на короткой шее, с мелким широким лицом. Обнаруживает тенденцию к ожирению.
Атлетик - имеет хорошую мускулатуру, крепкое телосложение, высокий или средний рост, широкий плечевой пояс и узкие бедра, выпуклые лицевые кости.
Система соматипирования Шелдона:
Чистый эндоморф характеризуется шарообразными формами, насколько это вообще возможно для человека. У такого индивидуума круглая голова, большой живот, слабые, вялые руки и ноги, с большим количеством жира на плечах и бедрах, но тонкие запястья и лодыжки. Подобного человека с большим количеством подкожного жира можно было бы назвать просто толстым, если бы все профильные размеры его тела (включая грудную клетку и таз) не превалировали над поперечными.ьЭтой конституции большой степени сопутствует избыточное жироотложение.
Чистый мезоморф - это классический Геркулес с преобладанием костей и мышц. У него массивная кубическая голова, широкие плечи и грудная клетка, мускулистые руки и ноги. Количество подкожного жира минимально, профильные размеры невелики.
Чистый эктоморф - это долговязый человек. У него худое, вытянутое лицо, сдвинутый назад подбородок, высокий лоб, узкая грудная клетка и живот, узкое сердце, тонкие и длинные руки и ноги. Подкожный жировой слой почти отсутствует, мускулатура неразвита. Явному эктоморфу совершенно не грозит ожирение.
2) Ротовая полость является начальным отделом пищеварительного тракта, где осуществляется анализ вкусовых свойств веществ и разделение их на пищевые и отвергаемые, защита пищеварительного тракта от попадания некачественных пищевых веществ и экзогенной микрофлоры, измельчение, смачивание слюной пищи, начальный гидролиз углеводов и формирование пищевого комка. Кроме того происходит раздражение механо-, хемо-, терморецепторов, вызывающее рефлекторное возбуждение деятельности слюнных желез, желез желудка, поджелудочной железы, печени, желез двенадцатиперстной кишки.
Слюна́ - прозрачная бесцветная жидкость, жидкая биологическая среда организма выделяемая в полость рта тремя парами крупных слюнных желез (подчелюстные, околоушные, подъязычные) и множеством мелких слюнными железами полости рта.
Слюна обладает pH от 5,6 до 7,6. На 98,5 % состоит из воды, содержит соли различных кислот, микроэлементы и катионы некоторых щелочных металлов, муцин (формирует и склеивает пищевой комок), лизоцим (бактерицидный агент), ферменты амилазу и мальтазу, расщепляющие углеводы до олиго- и моносахаридов, а также другие ферменты, некоторые витамины. Также состав секрета слюнных желез меняется в зависимости от характера раздражителя.
В среднем за сутки выделяется 1-2,5 л слюны. Слюноотделение находится под контролем вегетативной нервной системы. Центры слюноотделения располагаются в продолговатом мозге. Стимуляция парасимпатических окончаний вызывает образование большого количества слюны с низким содержанием белка. Наоборот, симпатическая стимуляция приводит к секреции малого количества вязкой слюны. Без стимуляции секреция слюны происходит со скоростью около 0,5 мл/мин.
Отделение слюны уменьшается при стрессе, испуге или обезвоживании и практически прекращается во время сна и наркоза. Усиление выделения слюны происходит при действии обонятельных и вкусовых стимулов, а также вследствие механического раздражения крупными частицами пищи и при жевании.
Жевание служит для механической переработки пищи, т.е. ее откусывания дробления, перетирания. При жевании пища смачивается слюной, и из нее формируется пищевой комок. Жевание происходит благодаря сложной координации сокращений мышц, обеспечивающих движения зубов, языка, щек, и дна полости рта.
Жевание сложнорефлекторный акт. Т.е. он осуществляется безусловно – и условно-рефлекторным механизмами. Безусловнорефлекторный состоит в том, что пищей раздражаются механорецепторы периодонта зубов и слизистой рта. От них импульсы по афферентным волокнам тройничного, языкоглоточного и верхнегортанного нервов поступают в центр жевания продолговатого мозга. По эфферентным волокнам тройничного, лицевого и подъязычного нервов импульсы идут к жевательным мышцам, осуществляя бессознательные согласованные сокращения. Условно-рефлекторные влияния позволяют произвольно регулировать жевательный акт.
Глотание - сложнорефлекторный акт, который начинается произвольно. Сформированный пищевой комок перемещается на спинку языка, языком прижимается к твердому небу и передвигается на корень языка. Здесь он раздражает механорецепторы корня языка и небных дужек. От них по афферентным нервам импульсы идут к центру глотания продолговатого мозга. От него, по эфферентным волокнам подъязычного, тройничного, языкоглоточного и блуждающего нерва, они поступают к мышцам полости рта, глотки, гортани, пищевода. Мягкое небо рефлекторно поднимается и закрывает вход в носоглотку. Одновременно гортань поднимается, а надгортанник опускается, закрывая вход в гортань. Пищевой комок проталкивается в расширившуюся глотку. Этим заканчивается ротоглоточная фаза глотания. Затем подтягивается пищевод и его верхний сфинктер расслабляется.
Начинается пищеводная фаза . По пищеводу пищевой комок продвигается за счет его перистальтики. Циркулярные мышцы пищевода сокращаются выше пищевого комка и расслабляются ниже него. Волна сокращения-расслабления распространяется к желудку. Этот процесс называется первичной перистальтикой. При подходе пищевого комка к желудку расслабляется нижний пищеводный или кардиальный сфинктер, пропуская комок в желудок. Вне глотания он закрыт и служит для предотвращения заброса в пищевод желудочного содержимого. Если пищевой комок застревает в пищеводе, то от места его расположения начинается вторичная перистальтика, по механизмам идентичная первичной. Твердая пища продвигается по пищеводу 8-9 сек. Жидкая стекает пассивно, без перистальтики, за 1-2 сек.
3) Гипо́физ - мозговой придаток в форме округлого образования, расположенный в турецком седле, вырабатывает гормоны, влияющие на рост, обмен веществ и репродуктивную функцию. Является центральным органом эндокринной системы; тесно взаимодействует с гипоталамусом.
Гормоны передней доли гипофиза
Адренокортикотропный гормон (АКТГ) - (адренокортикотропин, кортикотропин) - оказывает стимулирующее действие на функцию коркового вещества надпочечников.
Соматотропный гормон (СТГ) (гормон роста, соматотропин) - участвует в регуляции всех видов обмена веществ в организме человека, но основное действие - стимуляция роста скелета и увеличение размеров тела
Тиреотропный гормон (ТТГ) (тиреотропин) - биологическая роль состоит в поддержании нормального строения и функциональной активности щитовидной железы.
Лютеинизирующий гормон (ЛГ) (лютропин) - гонадотропный гормон, стимулирующий развитие половых желез у мужчин и женщин, а у женщин еще и овуляцию.
Пролактин (лактогенный гормон, лютеотропный гормон, лютеотропин, маммотропин) - обладает широкой биологической активностью: стимулирует рост и развитие молочных желез, рост и функцию сальных желез и рост внутренних органов. Пролактин стимулирует репродуктивные процессы, проявление материнского инстинкта, а у мужчин - рост предстательной железы.
Фолликулостимулирующий гормон (ФСГ) - стимулирует рост и развитие фолликулов в яичниках и сперматогенез в семенных пузырьках.
Гормоны задней доли гипофиза:
Окситоцин - гормон, стимулирующий отделение молока у женщин в период кормления и сокращение мускулатуры матки;
Вазопрессин (АДГ) - гормон, обладающий противодиуретическим и вазопрессорным действием, увеличивает реабсорбцию воды почкой, таким образом повышая концентрацию мочи и уменьшая её объём.
1) Возбудимость – это способность живой ткани отвечать на раздражение активной специфической реакцией – возбуждением, т.е. генерацией нервного импульса, сокращением, секрецией. Т.е. возбудимость характеризует специализированные ткани – нервную, мышечные, железистые, которые называются возбудимыми.
Возбуждение – это комплекс процессов реагирования возбудимой ткани на действие раздражителя, проявляющийся изменением мембранного потенциала, метаболизма и т.д. Возбудимые ткани обладают проводимостью. Это способность ткани проводить возбуждение. Наибольшей проводимостью обладают нервы и скелетные мышцы.
Раздражитель – это фактор внешней или внутренней среды действующий на живую ткань.
Хронаксиметрия - метод исследования возбудимости тканей в зависимости от времени действия раздражителя (определение хронаксии и реобазы).
Физиологическая лабильность (подвижность) – это большая или меньшая частота реакций, которыми может отвечать ткань на ритмическое раздражение. Чем быстрее восстанавливается ее возбудимость после очередного раздражения, тем выше ее лабильность. Определение лабильности предложено Н.Е. Введенским. Наибольшая лабильность у нервов, наименьшая у сердечной мышцы.
Порог раздражения – это минимальная сила раздражителя, при которой возникает возбуждение.
Пессимум - угнетение деятельности нервной и мышечной тканей, вызываемое чрезмерной частотой стимуляции нервного ствола, к-рая не может быть воспроизведена в виде биопотенциалов самого нерва и синхронных сокращений иннервируемой им мышцы. Ему соответствует такая частота, при к-рой каждое последующее раздражение падает на мышцу в фазу её абс. рефрактерности (невозбудимости). Пессимум сопровождается ослаблением сокращения мышцы в результате трансформации частоты раздражения.
Оптимум
- макс уровень деятельности нервной и
мышечной тканей, к-рый может быть
стабильно воспроизведён как в самом
нерве, так и в виде синхронных сокращений
иннервируемой им мышцы. Оптимум обусловлен
определённой частотой стимуляции
нервного ствола, при к-рой каждое
последующее раздражение поступает на
мышцу в фазу повышенной её возбудимости,
способствуя длительному слитному
сокращению - тетанусу.
2)Нервная регуляция сердечной деятельности осуществляется симпатическим и парасимпатическим отделами вегетативной нервной системы. Ядра блуждающего нерва, иннервирующего сердце, расположены в продолговатом мозге. Блуждающие нервы заканчиваются на интрамуральных ганглиях. Постганглионарные волокна правого вагуса идут к синоатриальному узлу, а левого к атриовентрикулярному. Кроме того они иннервируют миокард соответствующих предсердий. Парасимпатических окончаний в миокарде желудочков нет. Благодаря такой иннервации, правый вагус влияет преимущественно на частоту сердцебиений, а левый на скорость проведения возбуждения в атриовентрикулярном узле.
Тела симпатических нейронов, иннервирующих сердце, расположены в боковых рогах 5-ти верхних грудных сегментов спинного мозга. Аксоны этих нейронов идут к звездчатому ганглию. От него отходят постганглионарные волокна, многочисленные ветви которых иннервируют и предсердия и желудочки. В сердце имеется развитая внутрисердечная нервная система, включающая афферентные, эфферентные, вставочные нейроны и нервные сплетения. Ее считают отделом метасимпатической нервной системы.
Блуждающие нервы оказывают следующие воздействия на сердце:
1. Отрицательный хронотропный эффект. Это уменьшение частоты сердечных сокращений. Он связан с тем, что правый вагус тормозит генерацию импульсов в синоатриальном узле. Под действием вагуса их генерация может временно прекращаться.
2. Отрицательный инотропный эффект. Снижение силы сердечных сокращений. Обусловлен уменьшением амплитуды и длительности ПД, генерируемых клетками пейсмекерами.
3. Отрицательный дромотропный эффект. Понижение скорости проведения возбуждения по проводящей системе сердца. Связан с воздействием левого вагуса на атриовентрикулярный узел. При достаточно сильном его возбуждении возможно возникновение временной атриовентрикулярной блокады.
4. Отрицательный батмотропный эффект. Это уменьшение возбудимости сердечной мышцы. Под влиянием вагуса удлиняется рефрактерная фаза.
Эти воздействия вагусов на сердце обусловлены тем, что их окончания выделяют ацетилхолин. Он связывается с М-холинорецепторами кардиомиоцитов и вызывает гиперполяризацию их мембраны. Поэтому уменьшаются возбудимость, проводимость, автоматия кардиомиоцитов, а как следствие сила сокращений.
Если длительно раздражать блуждающие нервы, остановившееся первоначально сердце начинает вновь сокращаться. Это явление называется ускользанием сердца из под влияния вагуса. Оно является следствием параллельного усиления влияния симпатических нервов. Центры блуждающих нервов находятся в состоянии тонуса. Поэтому импульсы от них постоянно идут к сердцу.
Симпатические нервы противоположным образом воздействуют на сердечную деятельность. Они оказывают положительное хронотропное, инотропное, батмотропное и дромотропное влияния. Медиатор симпатических нервов норадреналин взаимодействует с b1-адренорецепторами мембраны кардиомиоцитов. Происходит ее деполяризация, а в результате ускоряется медленная диастолическая деполяризация в Р-клетках синоатриального узла, увеличиваются амплитуда и длительность генерируемых ПД, возрастает возбудимость клеток проводящей системы. Вследствие этого повышаются возбудимость, автоматия, проводимость и сила сокращений сердечной мышцы. Тонус симпатических центров регуляции сердечной деятельности выражен значительно слабее, чем парасимпатических.
3) Транспорт кислорода от легких к тканям осуществляется кровью в основном в виде химического соединения с гемоглобином - оксигемоглобина и в меньшей мере в растворенном состоянии.
Гемоглобин (Нb) это гемопротеин, содержащийся в эритроцитах. Молекулу гемоглобина образуют четыре субъединицы, каждая из которых включает гем, соединенный с атомом железом, и белковую часть глобин. Гем синтезируется в митохондриях эритробластов, а глобин в их рибосомах. У взрослого человека гемоглобин содержит две a – и две b-полипептидных цепи. Он называется А-гемоглобином (adult-взрослый). В зрелом возрасте он составляет основную часть гемоглобина.
Гем содержит атом 2-х валентного железа, который легко соединяется с кислородом и легко отдает его. При этом валентность железа не изменяется. Один грамм гемоглобина способен связывать 1,34 мл кислорода. Соединение гемоглобина с кислородом, образующееся в капиллярах легких называется оксигемоглобином (HbO2). Гемоглобин, отдавший кислород в капиллярах тканей, называется дезоксигемоглобином или восстановленным (Hb).
От 10 до 30% углекислого газа, поступающего из тканей в кровь, соединяются с амидной группировкой гемоглобина. Образуется легко диссоциирующее соединение карбгемоглобин (HbCO2). В этом виде часть углекислого газа транспортируется к легким.
В некоторых случаях гемоглобин образует патологические соединения. При отравлении угарным газом образуется карбоксигемоглобин (HbCO). Сродство гемоглобина с окисью углерода значительно выше, чем с кислородом, а скорость диссоциации карбоксигемоглобина в 200 раз меньше, чем оксигемоглобина. Поэтому присутствие в воздухе даже 1% угарного газа приводит к прогрессирующему увеличению количества карбоксигемоглобина и опасному угарному отравлению. Кровь теряет способность переносить кислород. При отравлении сильными окислителями, например нитритами, марганцевокислым калием, красной кровяной солью, образуется метгемоглобин (MetHb). В этом соединении гемоглобина железо становится трехвалентным. Поэтому метгемоглобин очень слабо диссоциирующее соединение. Он не отдает кислород тканям.
Кривая диссоциации оксигемоглобина:
В исходной ее точке, когда РаО2 гемоглобин не содержит кислорода и SaО2 также равняется нулю. По мере повышения Ра02 гемоглобин начинает быстро насыщаться кислородом, превращаясь в оксигемоглобин: небольшого увеличения напряжения кислорода оказывается достаточно для существенного прироста содержания НЬО2. При 40 мм рт. ст. содержание НЬО2 достигает уже 75 %. Затем наклон кривой становится все более и более пологим. На этом участке кривой гемоглобин уже менее охотно присоединяет к себе кислород, и для насыщения оставшихся 25 % НЬ требуется поднять Ра02 с 40 до 150 мм рт. ст. Впрочем, в естественных условиях гемоглобин артериальной Крови никогда не насыщается кислородом полностью, потому НТО при дыхании атмосферным воздухом Ра02 не превышает 100 мм рт. ст.
Кислородная емкость крови - количество кислорода, которое может быть связано кровью при её полном насыщении; выражается в объёмных процентах (об%); зависит от концентрации в крови гемоглобина. Определение Кислородной емкости крови важно для характеристики дыхательной функции крови. Кислородная емкость крови человека - около 18-20 об%.
1) Сон – это долговременное функциональное состояние, характеризующееся значительным снижением нервно – психической и двигательной активности, которое необходимо для восстановления способности мозга к аналитико – синтетической деятельности.
Виды сна:
1.Физиологический суточный сон.
2.Сезонный сон у животных (земляная белка 9 мес.)
3.Гипнотический сон.
4.Наркотический сон.
5.Патологический сон.
Продолжительность суточного сна у новорождённых около 20 часов, у годовалых детей 13-15 часов, у взрослых 6-9 часов. (Взгляды Наполеона на сон, вредная привычка, продолжительность жизни коротко спящих, средне спящих, длительно спящих людей).
В течение физиологического сна периодически друг друга сменяют 2 его формы: быстрый или парадоксальный сон, медленный сон. Быстрый сон возникает 4-5 раз за ночь и длится 1/4 всего времени сна. Во время быстрого сна мозг находится в длительном состоянии: об этом свидетельствует аa-ритм ЭЭГ, быстрые движения глазных яблок, подёргивание век, конечностей, учащаются пульс и дыхание и т.д. Если человека разбудить во время быстрого сна, он расскажет о сновидениях. При медленном сне этих явлений нет, а на ЭЭГ регистрируется дельта – ритм, свидетельствующий о тормозных процессах в мозге. Долгое время считалось, что во время медленного сна сновидений нет, сейчас установлено, что сновидения в этот период сна менее яркие, длительные и реальные. Возникновение ночных кошмаров также связано с медленным сном. Более того, обнаружено, что сомнабулизм или снохождение возникает именно во время медленного сна.
Значение сна:
1.Очищение Ц.Н.С. от метаболитов, накопившихся в процессе бодрствования.
2.Удаление накопившейся за день ненужной информации и подготовка к приёму новой.
3.Переход информации кратковременной памяти в долговременную. Он происходит во время медленного сна. Поэтому заучивание материала перед сном способствует запоминанию и лучшему воспроизведению запомненного. Особенно под улучшается запоминание логически несвязанного материала.
4.Эмоциональная перестройка. Во время быстрого сна происходит снижение возбудимости очагов мотивационного возбуждения, которые возникли в результате неудовлетворённой потребности. Во время сна неудовлетворённые потребности находят своё отражение в сновидениях (З. Фрейд. О сновидении). У больных с депрессивными состояниями наблюдаются необычайно яркие сновидения. Таким образом во сне происходит психологическая стабилизация и личность в определенной мере защищается от нерешённых конфликтов. Обнаружено, что люди мало спящие, у которых продолжительность быстрого сна относительно больше, лучше приспособлены к жизни и спокойно переживают психологические проблемы. Долго спящие обременены психологическими и социальными конфликтами.
2) В двенадцатиперстной кишке изливаются три вида пищеварительных соков: панкреатический (сок поджелудочной железы), желчь, кишечный сок. Все они имеют выраженную щелочную реакцию. В состав поджелудочного и кишечного сока входят три вида ферментов, расщепляющих белки, жиры и углеводы. Протеолитические ферменты: трипсин, химотрипсин, эластаза, карбоксипептидазы. Роль протеолитических ферментов заключается в распаде нативных белков и продуктов их первичной обработки в желудке (альбумоз и пептонов) до низкомолекулярных полипептидов и аминокислот. Амилолитические ферменты: альфа-амилаза. Их роль состоит в дальнейшем расщеплении углеводов до глюкозы и мальтозы. Липолитические ферменты: липаза, фосфолипаза А. Липаза секретируется в активном состоянии, ее активность возрастает под действием желчных кисло
Поджелудочная железа: Внешняя секреторная деятельность ее состоит в выделении в двенадцатиперстную кишку панкреатического сока, содержащего ферменты, участвующие в процессах пищеварения.
Регуляция образования и выделения панкреатического сока осуществляется гуморальным и нервным путями при участии секретина (гормона, образующегося при воздействии кислого желудочного содержимого в слизистой оболочке тонкой кишки) и секреторных волокон блуждающего и симпатического нервов. Физиологические стимуляторы отделения панкреатического сока- соляная и некоторые др. кислоты, жёлчь, пища. Состав панкреатического сока. В течение суток поджелудочная железа выделяет 1500-2000 мл сока. Панкреатический сок, полученный в чистом виде, представляет собой бесцветную прозрачную жидкость щелочной реакции (рН = 7,8- 8,4) благодаря наличию в ней двууглекислого натрия. Панкреатический сок содержит значительное количество плотных веществ (1,3%), что и обусловливает его высокий удельный вес (1,015). Из органических веществ в его состав входят главным образом белки, из неорганических - бикарбонаты, хлориды и другие соли. В панкреатическом соке содержатся также слизистые вещества, выделяемые железами выводного протока. Состав сока меняется в зависимости от того, вызвано ли его отделение раздражением блуждающего нерва или действием секретина. Но главной составной частью панкреатического сока являются ферменты, имеющие большое значение в процессах пищеварения. Эти ферменты следующие: трипсин, липаза, амилаза, мальтаза, пивертаза, лактаза, нуклеаза, а также в незначительном количестве эрепсин и ренин.
3) Терморегуляция это совокупность физиологических процессов теплообразования и теплоотдачи, обеспечивающих поддержание нормальной температуры тела. В основе терморегуляции лежит баланс этих процессов. Регуляция температуры тела посредством изменения интенсивности обмена веществ, называется химической терморегуляцией. Образование тепла усиливается путем интенсификации обменных процессов, это называется недрожательным термогенезом. Он обеспечивается за счет бурого жира. Его клетки содержат много митохондрий и специальный пептид, вызывающий разобщение процессов окисления и фософрилирования и стимулирующий распад липидов с выделением тепла. Кроме того термогенез усиливает непроизвольная мышечной активность в виде дрожи, произвольная моторной активность. Наиболее интенсивно теплообразование идет в работающих мышцах. При тяжелой физической работе оно возрастает на 500%.
Теплоотдача служит для выделения избытка образующегося тепла и называется физической терморегуляцией. Посредством теплоизлучения выделяется 60% тепла, конвекции (15%), теплопроводности (3 %), испарения воды с поверхности тела и из легких (20%).
Баланс процессов теплообразования и теплоотдачи обеспечивается нервными и гуморальными механизмами. При отклонении температуры тела от нормальной величины, возбуждаются терморецепторы кожи, сосудов, внутренних органов, верхних дыхательных путях. Этими рецепторами являются специализированные окончания дендритов сенсорных нейронов, а также тонкие волокна типа С. Холодовых рецепторов в коже больше, чем тепловых и они расположены более поверхностно. Нервные импульсы от этих нейронов по спиноталамическим трактам поступают в таламус, гипоталамус и кору больших полушарий. Формируется ощущение холода или тепла. В заднем гипоталамусе и преоптической области переднего, находится центр терморегуляции. Нейроны заднего гипоталамуса в основном обеспечивают химическую терморегуляцию, а переднего физическую. В центре имеется три типа нейронов. Первый термочувствительные нейроны. Они расположены в преоптической области и реагируют на изменение температуры крови, проходящей через мозг. Меньшее количество таких же нейронов имеется в спинном и продолговатом мозге. Вторая группа – интернейроны. Они получают информацию от периферических температурных рецепторов и терморецепторных нейронов. Эта группа нейронов служат для поддержания установочной точки, т.е. определенной температуры тела. Одна часть данных нейронов получает информацию от холодовых, другая от тепловых периферических рецепторов и терморецепторных нейронов. Третий тип нейронов – эфферентные. Они находятся в заднем гипоталамусе и обеспечивают регуляцию механизмов теплообразования.
Свои влияния на исполнительные механизмы, центр терморегуляции осуществляет через симпатическую и соматическую нервную системы, железы внутренней секреции. При повышении температуры тела возбуждаются периферические тепловые рецепторы и терморецепторные нейроны. Импульсы от них поступают к интернейронам, а затем к эффекторным. Эффекторными являются нейроны симпатических центров гипоталамуса. В результате их возбуждения активируются симпатические нервы, которые расширяют сосуды кожи и стимулируют потоотделение. При возбуждении холодовых рецепторов наблюдается обратная картина. Частота нервных импульсов идущих к кожным сосудам и потовым железам уменьшается, сосуды суживаются, потоотделение тормозится. Одновременно расширяются сосуды внутренних органов. Если это не приводит к восстановлению температурного гомеостаза, включаются другие механизмы. Во-первых, симпатические нервная система усиливает процессы катаболизма, а следовательно теплопродукцию. Выделяющийся из окончаний симпатических нервов норадреналин стимулирует процессы липолиза. Особую роль в этом играет бурый жир. Это явление называется не дрожательным термогенезом. Во-вторых, от нейронов заднего гипоталамуса начинают идти нервные импульсы к двигательным центрам среднего и продолговатого мозга. Они возбуждаются и активируют a-мотонейроны спинного мозга. Возникает непроизвольная мышечная активность в виде холодовой дрожи. Третий путь – это усиление произвольной двигательной активности. Большое значение имеет соответствующее изменение поведения, которое обеспечивается корой. Из гуморальных факторов наибольшее значение имеют адреналин, норадреналин и тиреоидные гормоны. Первые два гормона вызывают кратковременное повышение теплопродукции за счет усиления липолиза и гликолиза. При адаптации к длительному охлаждению усиливается синтез тироксина и трийодтиронина. Они значительно повышают энергетический обмен и теплопродукцию посредством увеличения количества ферментов в митохондриях.
1) Нерв – комплексное образование, состоящее из нервного волокна (миелинового или безмиелинового), рыхлой волокнистой соединительной ткани, образующей оболочку нерва.
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
Процессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием – с декрементом. Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «-». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70-120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
2) ПЕРЕДАЧА ВОЗБУЖДЕНИЯ В МИОКАРДЕ.
Появление электрических потенциалов в сердечной мышце связано с движение ионов через клеточную мембрану. Основную роль при этом играют катоины натрия и калия. Известно, что внутри клетки калия больше, чем в околоклеточной жидкости, концентрация внутриклеточного натрия, наоборот, меньше, чем околоклеточного. В состоянии покоя наружная поверхность клетки миокарда имеет положительный заряд в результате перевеса катионов натрия; внутренняя поверхность клеточной мембраны имеет отрицательный заряд в связи с перевесом внутри клетки анионов. В этих условиях клетка поляризована. Под влиянием внешнего электрического импулься клеточная мембрана становится проницаемой для катионов натрия, которые направляются внутрь клетки, и переносит туда свой положительный зарад. Наружная поверхность данного участка клетки приобретает отрицательный заряд в связи с перевесом там анаонов. Этот процесс называется ДЕПОЛЯРИЗАЦИЕЙ и связан с потенциалом действия. Скоро вся поверхность клетки снова приобретет отрицательный заряд, а внутренная – положительный. Таким образом, происходит ОБРАТНАЯ ПОЛЯРИЗАЦИЯ . Реполяризация мембраны вызывает постепенное закрывание калиевых и реактивацию натриевых каналов. В результате возбудимость миокардиальной клетки восстанавливается - это период так называемой относительной рефрактерности. В клетках рабочего миокарда (предсердия, желудочки) мембранный потенциал поддерживается на более или менее постоянном уровне.
Вышеперечисленные процессы происходит во времы систолы. Если вся поверхность снова приобретает положительный заряд, а внутренняя – отрицательный, то это соответствует диастоле. Во время диастолы происходит постепенные обратные движения ионов калия и натрия, которые мало влияют на заряд клетки, поскольку ионы натрия выходят из клетки, а ионы калия входят в нее одновременно. Эти процессы уравновешивают друг друга.
Вышенезванные процессы относятся к возбуждению единичного мышечного волокна миокарда. Возникнув при деполяризации, импульс вызывает возбуждение соседных участков миокарда, которые постепенно охватывает весь миокард, и развивается по типу цепной реакции. Возбуждение сердца начинается в снусном узле. Затем от синусного узла процесс возбуждения распространяется на предсердия. От предсердий оно идет к узлу. Обогнув это соединение, возбуждение переходит на ствол пучка Гиса.
Скорость распространения возбуждения различна в разных отделах проводящей системы, так в предсердиях и по пучку Гисса возбуждение распространяется со скоростью 1 м/с, по волокнам Пуркинье – 3 м/с, а в атриовентрикулярном узле со скоростью 0,05 м/с. Быстрое распространение возбуждение в предсердиях и желудочках обуславливает единовременный охват возбуждением всего миокарда. При этом его одновременное сокращение способствует увеличению силы выброса и эффективности работы. В то же время задержка возбуждения в атриовентрикулярном узле обеспечивает последовательное сокращение предсердий и желудочков, что также является очень важным моментом в гемодинамике.
ЭЛЕКТРОКАРДИОГРАФИЯ (ЭКГ) - является тестом, проведение которого позволяет получать ценную информацию о состоянии сердца. Суть данного метода состоит в регистрации электрических потенциалов, возникающих во время работы сердца и в их графическом отображении на дисплее или бумаге.
ПРИМЕНЕНИЕ
Определение частоты и регулярности сердечных сокращений (например, экстрасистолы (внеочередные сокращения), или выпадения отдельных сокращений - аритмии).
Показывает острое или хроническое повреждение миокарда (инфаркт миокарда, ишемия миокарда).
Может быть использована для выявления нарушений обмена калия, кальция, магния и других электролитов.
Выявление нарушений внутрисердечной проводимости (различные блокады).
Зубец Р отражает период возбуждения предсердий; зубец Q отражает период возбуждения межжелудочковой перегородки; зубец R самый высокий в ЭКГ, он соответствует периоду напряжения оснований желудочков; зубец S - полный охват миокарда желудочков возбуждением; зубец Т отражает полное восстановление мембранного потенциала клеток миокарда, т.е. потенциал покоя.
ЭКГ представляет собой запись суммарного электрического потенциала, появившегося при возбуждении множества миокардиальных клеток, а метод исследования называется электрокардиографией.
3) Половые железы (семенники у мужчин, яичники у женщин) относятся к железам со смешанной функцией, внутрисекреторная функция проявляется в образовании и секреции половых гормонов, которые непосредственно поступают в кровь.
Мужские половые гормоны – андрогены образуются в интерстициальных клетках семенников. Различают два вида андрогенов – тестостерон и андростерон .
Андрогены стимулируют рост и развитие полового аппарата, мужских половых признаков и появление половых рефлексов.
Они контролируют процесс созревания сперматозоидов, способствуют сохранению их двигательной активности, проявлению полового инстинкта и половых поведенческих реакций, увеличивают образование белка, особенно в мышцах, уменьшают содержание жира в организме. При недостаточном количестве андрогена в организме нарушаются процессы торможения в коре больших полушарий.
Женские половые гормоны эстрогены образуются в фолликулах яичника. Синтез эстрогенов осуществляется оболочкой фолликула, прогестерона – желтым телом яичника, которое развивается на месте лопнувшего фолликула.
Эстрогены стимулируют рост матки, влагалища, труб, вызывают разрастание эндометрия, способствуют развитию вторичных женских половых признаков, проявлению половых рефлексов, усиливают сократительную способность матки, повышают ее чувствительность к окситоцину, стимулируют рост и развитие молочных желез.
Прогестерон обеспечивает процесс нормального протекания беременности, способствует разрастанию слизистой эндометрия, имплантации оплодотворенной яйцеклетки в эндометрий, тормозит сократительную способность матки, уменьшает ее чувствительность к окситоцину, тормозит созревание и овуляцию фолликула за счет угнетения образования лютропина гипофиза.
Образование половых гормонов находится под влиянием гонадотропных гормонов гипофиза и пролактина. У мужчин гонадотропный гормон способствует созреванию сперматозоидов, у женщин – росту и развитию фолликула. Лютропин определяет выработку женских и мужских половых гормонов, овуляцию и образование желтого тела. Пролактин стимулирует выработку прогестерона.
Мелатонин тормозит деятельность половых желез.
Нервная система принимает участие в регуляции активности половых желез за счет образования в гипофизе гонадотропных гормонов. ЦНС регулирует протекание полового акта. При изменении функционального состояния ЦНС могут произойти нарушение полового цикла и даже его прекращение.
Менструальный цикл включает четыре периода.
1. Предовуляционный (с пятого по четырнадцатый день). Изменения обусловлены действием фоллитропина, в яичниках происходит усиленное образование эстрогенов, они стимулируют рост матки, разрастание слизистой оболочки и ее желез, ускоряется созревание фолликула, поверхность его разрывается, и из него выходит яйцеклетка – происходит овуляция.
2. Овуляционный (с пятнадцатого по двадцатьвосьмой день). Начинается с выхода яйцеклетки в трубу, сокращение гладкой мускулатуры трубы способствует продвижению ее к матке, здесь может произойти оплодотворение. Оплодотворенное яйцо, попадая в матку, прикрепляется к ее слизистой и наступает беременность. Если оплодотворение не произошло, наступает послеовуляционный период. На месте фолликула развивается желтое тело, оно вырабатывает прогестерон.
3. Послеовуляционный период. Неоплодотворенное яйцо, достигая матки, погибает. Прогестерон уменьшает образование фоллитропина и снижает продукцию эстрогенов. Изменения, возникшие в половых органах женщины исчезают. Параллельно уменьшается образование лютропина, что ведет к атрофии желтого тела. За счет уменьшения эстрогенов матка сокращается, происходит отторжение слизистой оболочки. В дальнейшем происходит ее регенерация.
4. Период покоя и послеовуляционный период продолжаются с первого по пятый день полового цикла.
Сперматогенез. Сперматогенез состоит из трех стадий и происходит в семенных канальцах мужских половых желез - семенниках (яичках). Первая стадия - многочисленный митоз спермообразу-ющих клеток; вторая - мейоз; третья - спермиогенез. Вначале образуются сперматогонии, расположенные на наружной стенке семенных канатиков. Затем они последовательно превращаются в сперматоциты первого порядка. Последние путем мейотического деления дают две одинаковые клетки - сперматоциты второго порядка. Во время второго деления сперматоциты второго порядка дают четыре незрелые половые клетки - гаметы. Они называются сперматидами. Образовавшиеся четыре сперматиды постепенно превращаются в активные движущиеся сперматозоиды.
1) Торможение условных рефлексов. В основе этого процесса лежат два механизма: безусловное (внешнее) и условное (внутреннее) торможение . И запредельное торможение. Безусловное торможение возникает мгновенно вследствие прекращения условно-рефлекторной деятельности. Выделяют внешнее и запредельное торможение.
Для активации внешнего торможения необходимо действие нового сильного раздражителя, способного создать в коре больших полушарий доминантный очаг возбуждения. В результате происходит торможение работы всех нервных центров, и временная нервная связь перестает функционировать. Такой вид торможения вызывает быстрое переключение на более важный биологический сигнал.
Запредельное торможение выполняет защитную роль и предохраняет нейроны от перевозбуждения, так как препятствует образованию связи при действии сверхсильного раздражителя.
Для возникновения условного торможения необходимо наличие специальных условий (например, отсутствие подкрепления сигнала). Различают четыре вида торможения:
1) угасательное (избавляет от ненужных рефлексов вследствие отсутствия их подкрепления);
2) дифферентное (приводит к сортировке близких раздражителей);
3) запаздывающее (возникает при увеличении продолжительности времени действия между двумя сигналами, приводит к избавлению от ненужных рефлексов, составляет основу для оценки уравновешенности и сбалансированности процессов возбуждения и торможения в ЦНС);
4) условный тормоз (проявляется только при действии дополнительного умеренного по силе раздражителя, который вызывает новый очаг возбуждения и тормозит остальные, является основой для процессов дрессировки и воспитания).
Торможение освобождает организм от ненужных рефлекторных связей и еще более усложняет отношения человека с окружающей средой. Запредельное торможение. Этот вид торможения отличается от внешнего и внутреннего по механизму возникновения и физиологическому значению. Оно возникает при чрезмерном увеличении силы или продолжительности действия условного раздражителя, вследствие того, что сила раздражителя превышает работоспособность корковых клеток. Это торможение имеет охранительное значение, так как препятствует истощению нервных клеток. По своему механизму оно напоминает явление "пессимума", которое было описано Н.Е.Введенским. Запредельное торможение может вызываться действием не только очень сильного раздражителя, но и действием небольшого по силе, но длительного и однообразного по характеру раздражения. Это раздражение, постоянно действуя на одни и те же корковые элементы, приводит их к истощению, а следовательно, сопровождается возникновением охранительного торможения. Запредельное торможение легче развивается при снижении работоспособности, например, после тяжелого инфекционного заболевания, стресса, чаще развивается у пожилых людей.
2) Клубочковая фильтрация . (~155-170л в сутки первичной мочи). Начальным этапом образвания мочи является фильтрация: в почечном тельце из капиллярного клубочка в полость капсулы фильтруется жидкая часть крови. Клубочковая фильтрация – это пассивный процесс. В условиях покоя у взрослого человека около 1 / 4 крови, выбрасываемой в аорту левым желудочком сердца, поступает в почечные артерии. Иными словами, через обе почки у взрослого мужчины проходит около 1300 мл крови в минуту, у женщин несколько меньше. Общая фильтрационная поверхность клубочков почек составляет примерно 1,5 м 2 . В клубочках из кровеносных капилляров в просвет капсулы почечного клубочка происходит ультрафильтрация плазмы крови, в результате чего образуется первичнаямоча, в которой практически отсутствует белок. В норме белки как коллоидные вещества не проходят через стенку капилляров в полость капсулы почечного клубочка. Клубочковая фильтрация, равна 100–125 мл за 1 мин. Суточное количество ультрафильтрата в 3 раза превышает общее количество жидкости, содержащейся в организме. Естественно, что первичная моча во время движения по почечным канальцам отдает большую часть своих составных частей, особенно воду, обратно в кровь. Лишь 1 % жидкости, профильтрованной клубочками, превращается в мочу.
В канальцах реабсорбируется 99% воды, натрия, хлора, гидрокарбоната, аминокислот, 93% калия, 45% мочевины и т.д. Из первичной мочи в результате реабсорбции образуется вторичная, или окончательная, моча, которая затем поступает в почечные чашки, лоханку и по мочеточникам попадает в мочевой пузырь.
Функциональное значение отдельных почечных канальцев в процессе мочеобразования неодинаково. Клетки проксимального сегмента нефрона реабсорбируют попавшие в фильтрат глюкозу, аминокислоты, витамины, электролиты; 6 / 7 жидкости, составляющей первичную мочу, подвергается реабсорбции также в проксимальных канальцах. Вода первичной мочи частично (парциально) реабсорбируется в дистальных канальцах. В этих же канальцах происходит дополнительная реабсорбция натрия, могут секрети-роваться в просвет нефрона ионы калия, аммония, водорода и др.
Регуляция СКФ осуществляется за счет нервных и гуморальных механизмов . Независимо от природы, регулирующие факторы влияют на СКФ за счет изменения: 1) тонуса артериол клубочков и, соответственно, объемого кровотока (плазмотока) через них и величины фильтрационного давления; 2) тонуса мезангиальных клеток и фильтрационной поверхности; 3) активности подоцитов и их «отсасывающей» функции.
С целью определения функции почечных клубочков на практике чаще всего используются методы определения скорости клубочковой фильтрации (СКФ) по клиренсу различных экзогенных и эндогенных веществ. Для вычисления количества жидкости, фильтрующейся в клубочках, используют физиологически инертное вещество, свободно проникающее через клубочковую мембрану с безбелковой частью плазмы. Соответственно его концентрация в клубочковой жидкости будет равной его концентрации в плазме крови. Если это вещество не реабсорбируется и не секретируется почечными канальцами, то оно будет выделяться с мочой в том же количестве, в котором прошло через клубочковый фильтр. Так как большая часть воды фильтрата подвергается обратному всасыванию, то вещество, используемое для определения объема фильтрата, сконцентрируется во столько раз, во сколько раз уменьшится объем воды в почечных канальцах. Клиренс любой субстанции вычисляют по формуле:
(1)C=(U×V)/P , где C - клиренс вещества (мл/мин), U - концентрация исследуемого вещества в моче (ммоль/л), Р - концентрация того же вещества в крови (ммоль/л), V - минутный диурез (мл/мин).
Для определения СКФ используются инулин, парааминогиппурат натрия, немеченный йогексол, (51)креатинин-этилендиаминтетрауксусная кислота ((51) Cr-ЭДТА). Оценка клубочковой фильтрации по клиренсу инулина признается «золотым стандартом» для определения почечной функции.
Дыхание - это совокупность процессов, благодаря которым организм потребляет кислород из окружающей среды и выделяет углекислый газ.
Этапы дыхания:
1. Внешнее дыхание /вентиляция легких/ - обмен газов между атмосферным воздухом и альвеолярным, легочная вентиляция.
2. Диффузия газов в легких - обмен газов между альвеолярным воздухом и кровью в капиллярах легких.
3. Транспорт газов кровью - этот этап осуществляется за счет деятельности сердечно-сосудистой системы, в результате чего кислород доставляется к тканям, а углекислый газ - к легким.
4. Диффузия газов в тканях - обмен газов между кровью и тканями.
5. Тканевое дыхание - окислительно-восстановительные реакции, протекающие с потреблением кислорода и выделением углекислого газа.
Первые 4 этапа изучает физиология, последний, 5-ый - биохимия.
Обеспечение тканей О2 и удаление из организма СО2 зависит от четырех процессов:
1.Вентиляция легких
2.Диффузия газов в альвеолы и ткани из крови и в кровь.
3.Перфузия легких кровью /интенсивность кровотока в легких/.
4.Перфузия тканей кровью
Отрицательное давление в плевральной щели играет важную роль в процессах вдоха и выдоха. Отрицательное давление в плевральной щели - это величина, на которую давление в плевральной щели ниже атмосферного; при спокойном дыхании оно равно 4 мм рт. ст. в конце выдоха и -8 мм рт. ст. в конце вдоха. Таким образом, реальное давление в плевральной щели составляет величину порядка 752756 мм рт. ст. и зависит от фазы дыхательного цикла. Отрицательное давление уменьшается в направлении сверху вниз примерно на 0,2 мм рт. ст. на каждый сантиметр, так как верхние отделы легких растянуты сильнее нижних, которые несколько сжаты под действием собственного веса.
Значение отрицательного давления в плевральной щели заключается в том, что оно 1) обеспечивает куполообразное положение диафрагмы, так как давление в грудной полости ниже атмосферного, а в брюшной полости оно несколько выше атмосферного за счет тонуса мышц стенки живота; 2) обеспечивает смещение диафрагмы вниз при сокращении ее мышцы во время вдоха; 3) способствует также притоку крови по венам к сердцу; 4) способствует сжатию грудной клетки при выдохе (см. п. 10.2 ниже).
Происхождение отрицательного давления. В процессе развития организма рост легких отстает от роста грудной клетки. Поскольку на легкое атмосферный воздух действует только с одной стороны - через воздухоносные пути, оно растянуто и прижато к внутренней стороне грудной клетки. Вследствие растянутого состояния легких возникает сила, стремящаяся вызвать спадение легких. Эта сила называется эластической тягой легких (ЭТЛ). О том, что легкие находятся в растянутом состоянии, свидетельствует факт их спадения при пневмотораксе (греч. рпе-ита - воздух, torax - грудь) - патологическом состоянии, возникающем при нарушении герметичности плевральной щели, в результате чего ее заполняет атмосферный воздух, оказываясь между висцеральным и париетальным листками плевры. Эластичность - способность ткани возвращаться в исходное состояние после прекращения действия растягивающей силы. Так как плевральная щель в норме не сообщается с атмосферой, давление в ней ниже атмосферного на величину ЭТЛ: при спокойном вдохе на 8мм рт. ст., при спокойном выдохе на 4ммрт. ст. Фильтрующаяся в плевральную щель жидкость всасывается обратно висцеральной и париетальной плеврами в лимфатическую систему, что является важным фактором в поддержании отрицательного давления в плевральной щели.
Составными элементами ЭТЛ являются: 1) эластиновые и кол-лагеновые волокна; 2) гладкие мышцы сосудов легких к, главное, 3) поверхностное натяжение пленки жидкости, покрывающей внутреннюю поверхность альвеол. Силы поверхностного натяжения составляют 2/3 величины ЭТЛ, причем величина поверхностного натяжения альвеолярной пленки существенно уменьшается в присутствии сурфактанта.
аким образом: Отрицательное давление в плевральной полости. Если измерить давление в плевральной полости во время дыхательной паузы, то можно обнаружить, что оно ниже атмосферного давления на 34 мм рт.ст., т.е. отрицательное. Это вызвано эластической тягой легких к корню, создающей некоторое разрежение в плевральной полости.
Во время вдоха давление в плевральной полости еще больше уменьшается за счет увеличения объема грудной клетки, а значит, отрицательное давление возрастает. Величина отрицательного давления в плевральной полости равна: к концу максимального выдоха - 1-2 мм рт. ст., к концу спокойного выдоха - 2-3 мм рт. ст., к концу спокойного вдоха -5-7 мм рт. ст., к концу максимального вдоха - 15-20 мм рт. ст.
Механизм вдоха. Вдох происходит с помощью трех одновременно протекающих процессов: 1) расширения грудной клетки; 2) увеличения объема легких; 3) поступления воздуха в легкие. У здоровых молодых мужчин разница между окружностью грудной клетки в положении вдоха и выдоха составляет 710 см, а у женщин 58 см.
Расширение грудной клетки при вдохе обеспечивается сокращением инспираторных мышц - диафрагмы, наружных межреберных и межхрящевых. Грудная клетка при вдохе расширяется в трех направлениях.
В вертикальном направлении грудная клетка расширяется в основном за счет сокращения диафрагмы и смещения ее сухожильного центра вниз, поскольку точки прикрепления периферических ее частей к внутренней поверхности грудной клетки по всему периметру находятся ниже купола диафрагмы. При спокойном вдохе купол диафрагмы опускается примерно на 2 см, при глубоком вдохе до 10 см. Диафрагмальная мышца главная дыхательная мышца, в норме вентиляция легких на 2/3 осуществляется за счет ее движений. Диафрагма принимает участие в обеспечении кашлевой реакции, рвоты, натуживания, икоты, в родовых схватках.
Во фронтальном направлении грудная клетка расширяется благодаря некоторому разворачиванию ребер в стороны при движении их вверх.
В сагиттальном направлении грудная клетка расширяется вследствие удаления концов ребер от грудины вперед при поднятии их.
Расширению грудной клетки способствуют также и силы ее упругости, так как грудная клетка в процессе выдоха сильно сжимается с помощью ЭТЛ, вследствие чего она стремится расшириться. Поэтому энергия при вдохе расходуется только на частичное преодоле ние ЭТЛ и брюшной стенки, а грудная клетка поднимается сама и расширяется при этом примерно до 60 % жизненной емкости. Спонтанно расширяющаяся грудная клетка способствует также преодолению ЭТЛ. Вместе с расширением грудной клетки расширяются и легкие. При расширении грудной клетки движение нижних ребер оказывает большее влияние на ее объем и вместе с движением диафрагмы вниз обеспечивает лучшую вентиляцию нижних долей легких, чем верхушек легких.
Увеличение объема легких при вдохе объясняют по-разному: легкие расширяются либо вследствие увеличения отрицательного давления в плевральной щели, либо силы адгезии (слипания париетального и висцерального листков плевры), либо того и другого.
По нашему мнению, легкие расширяются под действием атмосферного давления воздуха, направленного на них только с одной стороны (через воздухоносные пути); вспомогательную роль выполняют силы сцепления (адгезии) висцерального и париетального листков плевры. Сила, с которой легкие прижаты к внутренней поверхности грудной клетки атмосферным воздухом, равна Ратм.
С целью улучшения восприятия материала изменением величины давления в самих легких (на вдохе 2 мм рт. ст., на выдохе +2 мм рт. ст.) можно пренебречь.
Снаружи на грудную клетку действует Ратм, но на легкие оно не передается, поэтому на них действует только одностороннее атмосферное давление через воздухоносные пути. Поскольку снаружи на грудную клетку действует Ратм, а изнутри Ратм-Рэтл, при вдохе необходимо преодолеть силу ЭТЛ. Поскольку при вдохе ЭТЛ увеличивается вследствие расширения (растяжения)
легких, то увеличивается и отрицательное давление в плевральной щели. А это означает, что увеличение отрицательного давления в плевральной щели является не причиной, а следствием расширения легких.
Расширению легких при вдохе способствует сила сцепления (адгезии) между висцеральным и париетальным листком плевры. Но эта сила крайне мала по сравнению с атмосферным давлением, действующим на легкие через воздухоносные пути. Об этом свидетельствует тот факт, что легкие при открытом пневмотораксе спадаются, когда воздух поступает в плевральную щель и на легкие с обеих сторон (и со стороны альвеол, и со стороны плевральной щели) действует одинаковое атмосферное давление (см. рис. 10.2). Поскольку легкие в условиях пневмоторакса отрываются от внутренней поверхности грудной клетки, это означает, что ЭТЛ превосходит силу сцепления между париетальным и висцеральным листком плевры. Поэтому сила сцепления не может обеспечить растяжение легких при вдохе, так как она меньше ЭТЛ, действующей в противоположном направлении.
Все изложенное свидетельствует о том, что легкие следуют за расширяющейся грудной клеткой при вдохе, в основном вследствие действия на них атмосферного давления только с одной стороны - через воздухоносные пути. Оно действует постоянно - и на вдохе, и на выдохе. При расширении грудной клетки и легких давление в последних уменьшается примерно на 2 мм рт. ст., но такое уменьшение нельзя считать значительным, поскольку на легкие продолжает действовать давление, равное Ратм - 2 мм рт. ст. Это давление и прижимает легкие к внутренней поверхности грудной клетки - именно поэтому легкие следуют за расширяющейся грудной клеткой при вдохе.
Воздух поступает в легкие при их расширении вследствие некоторого (на 2 мм рт. ст.) падения давления в них. Этого незначительного градиента давления достаточно, поскольку воздухоносные пути имеют большой просвет и не оказывают существенного сопротивления движению воздуха. Кроме того, увеличение ЭТЛ при вдохе обеспечивает дополнительное расширение бронхов. Вслед за вдохом плавно начинается выдох, который при спокойном дыхании осуществляется без непосредственной затраты энергии.
Механизм выдоха. Выдох осуществляется вследствие одновременно происходящих трех процессов: 1) сужения грудной клетки; 2) уменьшения объема легких; 3) изгнания воздуха из легких. Экспираторными мышцами являются внутренние межреберные мышцы и мышцы брюшной стенки.
Сужение грудной клетки при выдохе обеспечивается ЭТЛ и эластической тягой брюшной стенки. Это достигается следующим образом. При вдохе растягиваются легкие, вследствие чего возрастает ЭТЛ. Кроме того, диафрагма опускается вниз и оттесняет органы брюшной полости, растягивая при этом саму брюшную стенку, вследствие чего увеличивается ее эластическая тяга. Как только прекращается поступление импульсов к мышцам вдоха по диафраг-мальным и межреберным нервам, прекращается возбуждение мышц вдоха, вследствие чего они расслабляются. После этого грудная клетка суживается под влиянием ЭТЛ и постоянно имеющегося тонуса мышц брюшной стенки при этом органы брюшной полости оказывают давление на диафрагму и поднимают ее.
Поднятию купола диафрагмы способствует также ЭТЛ. Сужению грудной клетки (опусканию ребер) способствует также ее масса, но главную роль играет ЭТЛ.
Механизм передачи ЭТЛ на грудную клетку и сужения ее. Это осуществляется за счет уменьшения давления атмосферного воздуха на грудную клетку изнутри через воздухоносные пути и легкие (см. рис. 10.2). Уменьшение давления равно силе ЭТЛ, так как с внутренней стороны реальное давление, оказываемое воздухом на грудную клетку, равно Pатм-Рэтл, а снаружи на грудную клетку действует Ратм Этот перепад давлений действует и на вдохе, и на выдохе, но вдоху он препятствует (преодоление ЭТЛ), а выдоху, наоборот, способствует. ЭТЛ сжимает грудную клетку, как пружину.
Сила сцепления (адгезии) висцерального и париетального листков плевры мала и не добавляется к ЭТЛ, и не вычитается из нее, а только способствует удержанию листков плевры друг с другом.
Легкие сжимаются при выдохе под действием их собственной эластической тяги, которая обеспечивает сужение и грудной клетки.
Воздух изгоняется из легких вследствие повышения давления в них (при спокойном выдохе - на 2 мм рт. ст.), так как объем легких при выдохе уменьшается, что ведет к сжатию воздуха и выдавливанию его из легких.
Дополнительно: При вдохе преодолевается ряд сил:
1) эластическое сопротивление грудной клетки,
2) эластическое сопротивление внутренних органов, оказывающих давление на диафрагму,
3) эластическое сопротивление легких,
4) вязко-динамическое сопротивление всех перечисленных выше тканей,
5) аэродинамическое сопротивление дыхательных путей,
6) силу тяжести грудной клетки,
7) силы инерции перемещаемых масс/органов/
Биомеханика спокойного вдоха и выдоха…
Биомеханика спокойного вдоха
В развитии спокойного вдоха играют роль: сокращение диафрагмы и сокращение наружных косых межреберных и межхрящевых мышц.
Под влиянием нервного сигнала диафрагма /наиболее сильная мышца вдоха/ сокращается, ее мышцы расположены радиально по отношению к сухожильному центру, поэтому купол диафрагмы уплощается на 1,5-2,0 см, при глубоком дыхании -на 10 см, растет давление в брюшной полости. Размер грудной клетки увеличивается в вертикальном размере.
Под влиянием нервного сигнала сокращаются наружные косые межреберные и межхрящевые мышцы. У мышечного волокна место прикрепления его к нижележащему ребру дальше от позвоночника, чем место его прикрепления к вышележащему ребру, поэтому момент силы нижележащего ребра при сокращении этой мышцы всегда больше, чем таковой у вышележащего ребра. Это приводит к тому, что ребра как бы приподнимаются, а грудные хрящевые концы как бы слегка скручиваются. Так как при выдохе грудные концы ребер располагаются ниже, чем позвоночные /дуга под углом/, то сокращение наружных межреберных мышц приводит их в более горизонтальное положение, окружность грудной клетки увеличивается, грудина приподнимается и выходит вперед, межреберное расстояние увеличивается. Грудная клетка не только приподнимается, но и увеличивает свои саггитальный и фронтальный размеры. За счет сокращения диафрагмы, наружных косых межреберных и межхрящевых мышц увеличивается объем грудной клетки. Движение диафрагмы обуславливает примерно 70-80% вентиляции легких.
Грудная клетка выстлана изнутри париетальным листком плевры, с которым крепко сращена. Легкое покрыто висцеральным листком плевры, с которым также крепко сращено. В нормальных условиях листки плевры плотно прилегают друг к другу и могут скользить /благодаря выделению слизи/ относительно друг друга. Силы сцепления между ними велики и листки плевры невозможно разъединить.
При вдохе париетальный листок плевры следует за расширяющейся грудной клеткой, тянет за собой висцеральный листок и тот растягивает ткань легкого, что приводит к увеличению их объема. В этих условиях воздух, находящийся в легких /альвеолах/ распределяется в новом, большем объеме, это приводит к падению давления в легких. Возникает разница давлений между окружающей средой и легкими /трансреспираторное давление/.
Трансреспираторное давление(Ртрр) - это разница между давлением в альвеолах (Ральв) и внешним /атмосферным/ давлением (Рвнеш). Ртрр= Ральв. - Рвнешн,. Равняется на вдохе - 4 мм рт. ст. Эта разница и заставляет войти порцию воздуха через воздухоносные пути в легкие. Это и есть вдох.
Биомеханика спокойного выдоха
Спокойных выдох осуществляется пассивно, т.е. не происходит сокращения мышц, а грудная клетка спадается за счет сил, которые возникли при вдохе.
Причины, вызывающие выдох:
1. Тяжесть грудной клетки. Поднятые ребра опускаются под действием тяжести.
2. Органы брюшной полости, оттесненные диафрагмой вниз при вдохе, поднимают диафрагму.
В дыхательных движениях участвуют:
1. Дыхательные пути, которые по своим свойствам являются слегка растяжимыми, сжимаемыми и создают поток воздуха.
К воздухоносным путям, управляющим потоком воздуха, относятся: нос, полость носа, носоглотка, гортань, трахея, бронхи и бронхиолы.
Нос и полость носа служат проводящими каналами для воздуха, где он нагревается, увлажняется и фильтруется.
Полость носа выстлана богато васкулиризированной слизистой оболочкой. В верхней части полости носа лежат обонятельные рецепторы. Носовые ходы открываются в носоглотку.
Гортань лежит между трахеей и корнем языка.
У нижнего конца гортани начинается трахея и спускается в грудную полость, где делится на правый и левый бронхи.
Дыхательные пути от трахеи до концевых дыхательных единиц (альвеол) ветвятся (раздваиваются) 23 раза.
Первые 16 «поколений» дыхательных путей – бронхи и бронхиолы выполняют проводящую функцию.
«Поколения» 17...22, респираторные бронхиолы и альвеолярные ходы, составляют переходную (транзиторную) зону.
И только 23-е «поколение» является дыхательной респираторной зоной и целиком состоит из альвеолярных мешочков с альвеолами.
Общая площадь поперечного сечения дыхательных путей по мере ветвления возрастает более чем в 4,5 тысячи раз. Правый бронх обычно короче и шире левого.
2. Эластическая и растяжимая легочная ткань.
Легкие состоят из бронхиол и альвеолярных мешочков, а также из артерий, капилляров и вен легочного круга кровообращения.
Респираторный отдел представлен альвеолами.
В легких имеется три типа альвеолоцитов (пневмоцитов), выполняющих разную функцию.
Альвеолоциты второго типа осуществляют синтез липидов и фосфолипидов легочного сурфактанта.
Общая площадь альвеол у взрослого человека достигает 80...90 м2, т.е. примерно в 50 раз превышает поверхность тела человека.
3. Грудная клетка, состоящая из пассивной костно-хрящевой основы, которая соединена соединительными связками и дыхательными мышцами, которые осуществляют поднятие и опускание ребер и движения купола диафрагмы.
За счет большого количества эластической ткани легкие, обладая значительной растяжимостью и эластичностью, пассивно следуют за всеми изменениями конфигурации и объема грудной клетки.
Существуют два механизма, вызывающие изменение объема грудной клетки: поднятие и опускание ребер и движения купола диафрагмы.
Дыхательные мышцы подразделяются на инспираторные и экспираторные.
Инспираторными мышцами являются диафрагма, наружные межреберные и межхрящевые мышцы.
При спокойном дыхании объем грудной клетки изменяется в основном за счет сокращения диафрагмы и перемещения ее купола.
Опусканию диафрагмы всего на 1 см соответствует увеличение емкости грудной полости примерно на 200...300 мл.
При глубоком форсированном дыхании участвуют дополнительные мышцы вдоха: трапециевидные, пепередние лестничные и грудино-ключично-сосцевидные мышцы.
Они включаются в активный процесс дыхания при значительно больших величинах легочной вентиляции, например, при восхождении альпинистов на большие высоты или при дыхательной недостаточности, когда в процесс дыхания вступают почти все мышцы туловища.
Экспираторными мышцами являются внутренние межреберные и мышцы брюшной стенки, или мышцы живота.
Каждое ребро способно вращаться вокруг оси, проходящей через две точки подвижного соединения с телом и поперечным отростком соответствующего позвонка.
Верхние отделы грудной клетки на вдохе расширяются преимущественно в переднезаднем направлении, а нижние отделы больше расширяются в боковых направлениях, так как ось вращения нижних ребер занимает сагиттальное положение.
В фазу вдоха наружные межреберные мышцы, сокращаясь, поднимают ребра, а в фазу выдоха ребра опускаются благодаря активности внутренних межреберных мышц.
При обычном спокойном дыхании выдох осуществляется пассивно, поскольку грудная клетка и легкие спадаются - стремятся занять после вдоха то положение, из которого они были выведены сокращением дыхательных мышц.
Однако при кашле, рвоте, натуживании мышцы выдоха активны.
При спокойном вдохе увеличение объема грудной клетки составляет примерно 500...600 мл.
Движение диафрагмы во время дыхания обусловливает до 80% вентиляции легких.
Дыханием называется комплекс физиологических процессов, обеспечивающих обмен кислорода и углекислого газа между клетками организма и внешней средой. Оно включает следующие этапы:
1. Внешнее дыхание или вентиляция. Это обмен дыхательных газов между атмосферным воздухом и альвеолами.
2. Диффузия газов в легких, т.е. их обмен между воздухом альвеол и кровью.
3. Транспорт газов кровью.
4. Диффузия газов в тканях. Обмен газов между кровью и внутриклеточной жидкостью.
5. Клеточное дыхание. Поглощение кислорода и образование углекислого газа в клетках.
Механизм внешнего дыхания.
Внешнее дыхание осуществляется в результате ритмических движений грудной клетки. Дыхательный цикл состоит из фаз вдоха (inspiratio) и выдоха (exspiratio), между которыми отсутствует пауза. В покое у взрослого человека частота дыхательных движений 16-20 в минуту. Вдох это активный процесс. При спокойном вдохе сокращаются наружные межреберные и межхрящевые мышцы. Они приподнимают ребра, а грудина отодвигается вперед. Это ведет к увеличению сагиттального и фронтального размеров грудной полости. Одновременно сокращаются мышцы диафрагмы. Ее купол опускается, и органы брюшной полости сдвигается вниз, в стороны и вперед. За счет этого грудная полость увеличивается и в вертикальном направлении. После окончания вдоха дыхательные мышцы расслабляются. Начинается выдох. Спокойный выдох – пассивный процесс. Во время него происходит возвращение грудной клетки в исходное состояние. Это происходит под действием ее собственного веса, натянутого связочного аппарата и давления на диафрагму органов брюшной полости. При физической нагрузке, патологических состояниях, сопровождающихся одышкой (туберкулез легких, бронхиальная астма и т.д.) возникает форсированное дыхание. В акт вдоха и выдоха вовлекаются вспомогательные мышцы. При форсированном вдохе дополнительно сокращаются грудино-ключично-сосцевидные, лестничные, грудные и трапециевидные мышцы. Они способствуют дополнительному поднятию ребер. При форсированном выдохе сокращаются внутренние межреберные мышцы, которые усиливают опускание ребер, т.е. это активный процесс. Различают грудной и брюшной типы дыхания. При первом дыхание в основном осуществляется за счет межреберных мышц, при втором – за счет мышц диафрагмы. Грудной или реберный тип дыхания характерен для женщин. Брюшной или диафрагмальный – для мужчин. Физиологически более выгоден брюшной тип, т.к. он осуществляется с меньшей затратой энергии. Кроме того, движения органов брюшной полости при дыхании препятствует их воспалительным заболеваниям. Иногда встречается смешанный тип дыхания.
Несмотря на то, что легкие не сращены с грудной стенкой, они повторяют ее движения. Это объясняется тем, что между ними имеется замкнутая плевральная щель. Изнутри стенка грудной полости покрыта париетальным листком плевры, а легкие ее висцеральным листком. В межплевральной щели находится небольшое количество серозной жидкости. При вдохе объем грудной полости возрастает, а так как плевральная полость изолирована от атмосферы, то давление в ней понижается. Легкие расширяются, давление в альвеолах становится ниже атмосферного. Воздух через трахею и бронхи поступает в альвеолы. Во время выдоха объем грудной клетки уменьшается. Давление в плевральной щели возрастает, воздух выходит из альвеол. Движения или экскурсии легких обеспечиваются колебаниями отрицательного межплеврального давления. После спокойного выдоха оно ниже атмосферного на 4-6 мм рт ст. На высоте спокойного вдоха на 8-9 мм рт ст. После форсированного выдоха оно ниже на 1-3 мм рт ст., а форсированного вдоха на 10-15 мм рт ст. Наличие отрицательного межплеврального давления объясняется эластической тягой легких. Это сила, с которой легкие стремятся сжаться к корням, противодействуя атмосферному давлению. Она обусловлена упругостью легочной ткани, которая содержит много эластических волокон. Кроме того, эластическую тягу увеличивает поверхностное натяжение альвеол. Изнутри они покрыты пленкой сурфактанта. Это липопротеид, вырабатываемый митохондриями альвеолярного эпителия. Благодаря особому строению его молекул, на вдохе он повышает поверхностное натяжение альвеол, а на выдохе, когда их размеры уменьшаются, наоборот понижает. Это препятствует спаданию альвеол, т.е. возникновению ателектаза. При генетической патологии, у некоторых новорожденных нарушается выработка сурфактанта. Возникает ателектаз и ребенок гибнет. В старости, а также при некоторых хронических заболеваниях легких, количество эластических волокон возрастает. Это явление называется пневмофиброзом. Дыхательные экскурсии затрудняются. При эмфиземе эластические волокна наоборот разрушаются, и эластическая тяга легких снижается. Альвеолы раздуваются, величина экскурсии легких также уменьшается. При попадании воздуха в плевральную полость возникает пневмоторакс. Различают его следующие виды:
1. По механизму возникновения: патологический (рак легких, абсцесс, проникающее ранение грудной клетки) и искусственный (лечение туберкулеза).
2. В зависимости от того, какой листок плевры поврежден, выделяют наружный и внутренний пневмоторакс.
3. По степени сообщения с атмосферой различают открытый пневмоторакс, когда плевральная полость постоянно сообщается с атмосферой. Закрытый, если произошло однократное попадание воздуха. Клапанный, когда на вдохе воздух из атмосферы входит в плевральную щель, а на выдохе отверстие закрывается.
4. В зависимости от стороны повреждения – односторонний (правосторонний, левосторонний), двусторонний.
Пневмоторакс является опасным для жизни осложнением. В результате него легкое спадается и выключается из дыхания. Особенно опасен клапанный пневмоторакс.
Билет 22
23. Гормональная регуляция обмена кальция в организме. Паратиреоидные гормоны, кальцитонин, кальцитриол, их функции
Одновременно с механизмом, обеспечиваемым существованием обмениваемых солей кальция в костях, который работает как буферная система по отношению к концентрации ионов кальция в межклеточной жидкости, оба гормона (паратгормон и кальцитонин) начинают действовать в течение 3-5 мин после быстрого изменения концентрации ионов кальция. Скорость секреции ПТГ возрастает; как уже объяснялось, это запускает в действие многочисленные механизмы, направленные на снижение концентрации ионов кальция. Одновременно со снижением концентрации ПТГ концентрация кальцитонина возрастает у молодых животных и, вероятно, у маленьких детей (и у взрослых, но в меньшей степени). Кальцитонин вызывает быстрое поступление кальция в кости, а также, возможно, и во многие клетки других тканей, поэтому у очень молодых животных избыток кальцитонина может быть причиной того, что высокая концентрация ионов кальция возвращается к норме значительно быстрее, чем это может осуществить в одиночку буферная система, опосредованная механизмом легкообмениваемых солей кальция. В случае продолжительно существующего избытка кальция или его дефицита только влияния ПТГ оказываются действительно важными в нормализации концентрации ионов кальция в плазме. В случае длительного дефицита кальция в рационе ПТГ часто может стимулировать выход кальция из костей в количествах, достаточных для поддержания нормальной его концентрации в плазме в течение одного года, но очевидно, что даже этот источник кальция может иссякнуть. По обнаруживаемому эффекту кости можно считать буферным резервом кальция, которым управляет паратгормон. Если кости в качестве источника кальция иссякнут либо, напротив, переполнятся кальцием, в роли долговременного механизма, управляющего концентрацией кальция во внеклеточной жидкости, выступит ПТГ и витамин D, регулирующие всасывание кальция в кишечнике и его экскрецию с мочой.
Если паратиреоидные железы не секретируют достаточное количество паратгормона, это приводит к снижению вымывания остеоцитами легкообмениваемого кальция из костей при почти полной и повсеместной инактивации остеокластов. В результате абсорбция кальция из костей настолько уменьшается, что это приводит к снижению уровня кальция в жидких средах организма. Вследствие того, что кальций и фосфаты перестают вымываться из костей, кости обычно остаются прочными.
Кальцитонин
- пептидный гормон, состоящий из 12 аминокислот, физиологическая функция которого заключается в регуляции обмена кальция и фосфора. Интерес к этому гормону объясняется, в первую очередь, его участием в обеспечении относительно постоянного уровня кальция.
Основным и непосредственным фактором, который действует на щитовидную железу и активизирует синтез выделения кальцитонина. есть концентрация кальция в сыворотке кропи. Повышение уровня кальция в крови, особенно его ионизированной формы, усиливает секрецию кальцитонина, а снижение - угнетает.
Опосредованный путь регуляции секреции кальцитонина связан с секрецией гастрина и некоторых других энторогормонов. Уменьшение уровня кальция в пищеварительном тракте способствует секреции гастрина, что, в свою очередь, приводит к усилению синтеза и выделения кальцитонина щитовидной железой.
Кальцитонин через специфические рецепторы (в костях, почках) воздействует на цАМФ. В результате, прежде всего, тормозится резорбция костей и стимулируется их минерализацию.в частности, проявляется понижением уровня кальция и фосфора в сыворотке крови и экскреции гидроксипролина.
Паратиреоидный гормон (ПГТ)
является функциональным антагонистом кальцитонина: первый обеспечивает увеличение состава кальция, а второй - его снижение. Низкая концентрация кальция в плазме крови стимулирует поступление в кровь значительного количества ПТГ. который увеличивает реабсорбцию кальция в канальцах почек и секрецию фосфатов, а в костной ткани - ускорение процесса резорбции и освобождения кальция в межклеточное пространство.
На уровне клеток кальцитонин влияет на транспорт кальция через ее мембрану. Он стимулирует поглощение кальция митохондриями и тем самым задерживает отток кальция из клеток. Этот процесс связан с активностью аденозинтрифосфорной кислоты (АТФ) клеточной мембраны и зависит от соотношения натрия и калия. Кальцитонин влияет на органический состав кости: угнетает распад коллагена, что проявляется уменьшением экскреции с мочой оксипролина